
|
Improvement of nitrate and nitrite reduction rates prediction Carolina Estuardo M. Cristina Martí César Huiliñir Estrella Aspé Lillo Marlene Roeckel von Bennewitz* *Corresponding author Financial support: Funded by grant FONDECYT Nº 1040495 (Chile). Keywords: denitrification kinetics, nitrate reduction, nitrite reduction, pH.
Reported models of denitrification rates integrate in an unique parameter the pH-dependent inhibition by HNO2 and the pH effect on the bacterial metabolic activity; furthermore, they do not quantify separately the pH effect on the nitrate and on the nitrite reduction rates. The goal of this work was to quantify both effects on the kinetics of nitrate and nitrite reduction to improve the models’ predictive value. Assays were performed at a pH range of 6.5-9.0 in batch reactors at 37ºC with an activated sludge. At the studied pH range and at below the HNO2 inhibitory concentration (0.004 mg L-1), the maximum nitrate reduction rate diminished 23% and 50% by decreasing or increasing, respectively, one pH unit from 8.0. The maximum nitrite reduction at pH 8.0 diminished 15% at pH 7.0 and 40% at pH 9.0. At HNO2 concentrations over the inhibitory concentration, except at pH > 8.0, the maximum nitrate reduction rate diminished 50% upon decreasing the pH from 8.0 to 7.0 or increasing it from 8.0 to 9.0. Inclusion of the pH effect in the reported models improved their predictive value; average deviations from the experimental data were reduced from 53% to 10.7% or 33.8% to 10.5% for nitrite and nitrate reduction rates, respectively.
Several food processing industries discharge their liquid effluents with high organic load to the environment causing ecological and public health problems. Salmon-plant effluents have high protein (4.08- 22.5 g COD L-1) and salinity loads (River et al. 1998). The salinity of these effluents (24 g Na Cl L-1) derives from the utilization of seawater (approximately 30 g Na Cl L-1) in some of their processing. Due to their high carbon load, any biological treatment of these effluents should start by an anaerobic degradation. The anaerobic treatment of protein gives rise to ammonia; thus, although the original nitrate content in these effluents is almost negligible, further steps are required to aerobically oxidize the ammonia to nitrate (nitrification) and anoxically reduce nitrate to gaseous nitrogen (denitrification). Several authors have suggested that HNO2 accumulation inhibit denitrification (Glass and Silverstein, 1998; Soto et al. 2007). Nitrite is an intermediate in the denitrification process and depending on the pH and temperature of the medium could prevail as the ionized or the non-ionized form. The pH can affect directly the bacterial growth and its enzymatic activities (Campos and Flotats, 2003), including denitrifying enzymes, and indirectly affect the denitrification rate through changes in the concentration of HNO2. If the non-ionized form of nitrous acid prevails, it can mask the direct inhibitory effect of the pH. Almeida et al. (1995) and Wild et al. (1995) have developed kinetic models to predict denitrification rates. However, these models have considered the apparent net pH effect, i.e., the pH-dependent inhibitory effect of HNO2 and the pH effect on the bacteria activity in their kinetic expression. Moreover, Almeida et al. (1995) worked with a pure culture of P. fluorescens at 28ºC while Wild et al. (1995) reported kinetic parameters at 20ºC. Anoxic activated sludges usually include Pseudomonas fluorescens, Pseudomonas denitrificans, Paracoccus denitrificans and Micrococcus denitrificans, between other bacteria. A mixed bacterial population, as the ones used for effluents treatment, could exhibit different sensibilities to pH changes in the medium and to inhibitors than the ones reported in pure cultures. The rate of nitrate reduction has been represented as a function of the nitrate concentration by the Monod equation (Almeida et al. 1995) and the rate of nitrite reduction has been represented by a Haldane-type kinetics, since high HNO2 concentrations inhibits this rate (Almeida et al. 1995; Glass and Silverstein, 1998). Although it has been reported that the denitrification rate varies with a change in the pH (Almeida et al. 1995; Glass and Silverstein, 1998), the quantification of the direct pH effect on this rate or its inclusion in the kinetics of denitrification have not been reported in the literature. There are different expressions for the pH-inhibition function for biological processes reported in the literature. Ramsay and Pullammanapallil (2005) modelled the effect of the pH, up to pH 7.0, on the acidogenic bacteria by an empiric relationship of the exponential type. Siegrist et al. (2002) included a non-competitive quadratic inhibition factor due to pH inhibition up to pH 7.0. Since the former relationships are empiric ones and for pH values below 7.0, the Michaelis function (Segel, 1975), initially proposed to quantify the dependence of the enzymatic activity on the pH, seems the more appropriate expression to model the effect of the pH on the denitrification rate. This function has been modified to quantify the inhibitory effect of a wide range of pH values on the acetogenic step of the anaerobic process (Angelidaki et al. 1993; Batstone et al. 2002). The goal of this work was to include the effect of the pH on the kinetics of nitrate reduction and nitrite reduction in the modelling of these rates, and evaluate its predictive value using experimental data obtained with a mixed denitrifying sludge for treatment at 37ºC of a salmon plant effluent. Validation of the inclusion of the pH effect in the kinetics of denitrification will be assessed by calculating the fitting deviations of the reported models with and without the inclusion of pH function. An improvement of the predictive values of these models will broaden the use of the kinetics to different operational conditions. An adapted, stable biomass, able to carry out denitrification and methanogenesis was obtained from 3 L anaerobic and anoxic reactors that were at steady state for at least a year. These reactors were fed with 50% of a salmon-plant effluent and 50% of a synthetic substrate (v/v); the latter to adapt the sludge to nitrate. The average industrial effluent composition was (g L-1): NO3-, 0; NO2-; 0; NH4+, 0.023; total Kjeldahl nitrogen, 10.755; TOC, 2.482; COD, 6.469; NaCL, 24; protein, 2.536; SO4=, 0.001. The synthetic substrate composition was (g L-1): KH2PO4, 3; K2HPO4, 3; NaHCO3, 0.4; MgSO4·7H2O, 0.005; NaCl, 24; yeast extract, 0.05; peptone casein, 0.12; KNO3, 0.721; FeSO4·7H2O, 0.0011; CaCl2, 0.0005; 17.5 M acetic acid, 1.22 mL4. Kinetic assays at constant pH were carried out in 1-L batch reactors with a useful volume of 800 mL of anoxic medium. They were seeded with 20% (v/v) of the adapted inoculum (50% anaerobic and 50% anoxic); the purpose of seeding the reactors with a large microbial concentration was to ensure a constant biomass throughout the assays. The reactors were fed with a mixture (v/v) of 50% salmon plant effluent and 50% of a synthetic substrate of the following composition (g L-1): NaHCO3, 0.4; MgSO4 7H2O, 0.005; NaCl, 24; yeast extract, 0.05; peptone casein, 0.12; FeSO4·7H2O, 0.0011; CaCl2, 0.0005; KCl, 0.0005 CoCl2, 0.0005; 17.5 M acetic acid, 1.22 mL. In assays performed with initial nitrate concentrations of 10 or 80 mg NO3- -N L-1, nitrate was added to the medium as KNO3. A C/N of 10 was used to avoid dissimilatory degradation of nitrate and nitrite to ammonia. Anoxic conditions in the reactors were obtained by gassing for approximately 1 min with N2 and sealing the reactors with butyl stoppers. Temperature was kept constant at 37 ± 1ºC by means of a thermostatic bath; this assay temperature was chosen as it is the optimum temperature for anaerobic digestion. The pH was controlled (± 0.1 units) by a Cole Palmer (ORP 5595, USA) controller; the pH was adjusted by the addition of 1 N HCl or 1 N NaOH. Samples were withdrawn every 40 min for 12 hrs and every 2 hrs for 16 hrs in kinetic assays performed at an initial nitrate concentration of 10 and 80 mg NO3--N L-1, respectively. Liquid samples were centrifuged at 3500 rpm for 4 min and filtered under vacuum with a 0.45 µm filter membrane for bacteria removal. In each sample, nitrite, nitrate, total ammonia nitrogen (TAN), volatile suspended solids (VSS) and total organic carbon (TOC) were measured as described elsewhere (Aspé et al. 2001; Sánchez et al. 2005). VSS were measured at the beginning and at the end of each assay. Total ammonia nitrogen was measured according to Standard Methods (APHA, 1992). Analyses were performed in triplicate. Maximal rates of nitrate and nitrite reduction.The kinetic analysis used a simplified denitrification model:
Although other reduced products of denitrification have been reported, pH was thought to affect specially the nitrate and nitrite reduction rates. The nitrate consumption rate can be described by:
Where r1 is
the rate of disappearance of nitrate, k1 is
a rate constant and The rate of nitrite consumption is:
Where Nitrite is simultaneously formed and consumed, thus: rNet nitrite = -r1 + r2 [4] The initial maximal rates of nitrate consumption (r1) were obtained from the slope of a nitrate concentration vs time plot at different pHs. The net rate between nitrite accumulation and nitrite consumption, i.e. rnet nitrite, was obtained from a nitrite concentration vs. time plot at different pHs and measuring the slope after nitrite has reached its maximal accumulation. Modelling of the pH effectSince the sludge used to denitrify in the present work is an anaerobic sludge, it is feasible to quantify the inhibitory effect of the pH on the denitrifying bacteria through mathematical functions representing the effect of the pH on anaerobic (acidogenic) bacteria. Several studies have included the inhibitory effect of the pH on the acidogenic bacteria as a factor that multiplies the substrate consumption rate: ri = f(Ci)·f(pH) [5] Where ri: Degradation rate of the compound i. f(Ci): A mathematical expression that describes the kinetics of the substrate i degradation as a function of the substrate concentration used. f(pH): A mathematical expression that quantifies the pH effect. The Michaelis function used to model the effect of the pH on enzymatic reactions was used to model the effect of the pH on nitrate and nitrite consumption rate (Segel, 1975):
Or: where: r, is the maximum apparent rate; rmax, is the maximum rate at the optimum pH; [H+], is the proton concentration; Ks1 is the lowest proton concentration where r is equal to ½rmax; Ks2 is the highest proton concentration where r is equal to ½rmax; pKs1, is the logarithm of the lowest pH at which r is equal to ½rmax; pKs2 is the logarithm of the highest pH at which r is equal to ½rmax. If the pK values at which r is equal to ½rmax are separated by more than 3.5 pH units the model should fit 99% the data. On the contrary, if they are separated by less than 3.5 pH units, the maximum of the plot rmax vs pH will occur at a value significantly lower than the theoretical maximum and, consequently, the pH values at the half-maximum points will not correspond to the pK values (Segel, 1975) and, thus, the Michaelis pH function must be modified by an empiric factor (A) so that it reaches rmax as a central value (Glass et al. 1997). Therefore, equation (7) assumes the following form: The concentration vs time plots obtained from kinetic parameters reported in the literature were made integrating mass balances in batch reactor on software Matlab v 6.5. The pKs1 and pKs2 values were obtained by fitting of the r/rmax vs pH plot by equation (8). The deviation between the experimental data and the values given by the model using kinetic parameters reported in the literature, were calculated considering the experimental data as the true values.
Effect of the pH on the apparent maximum specific rate of nitrate reduction Assays at different pH values and different nitrate concentrations were carried out to determine the inhibitory effect of the pH on denitrification. Figure 1a and Figure 1b shows the nitrate and nitrite concentration variation in assays performed at pH 8.0 and an initial nitrate concentration of 10 mg NO3--N L-1 and 80 mg NO3--N L-1, respectively. As shown, nitrate concentration diminished with time while the nitrite concentration increased up to a peak at approximately 5 hrs of starting the assay. A similar behaviour was observed at pH values of 6.5, 7.0, 8.0, 8.5 and 9.0. In these assays, TAN concentration varied between 64 and 142 mg TAN L-1 and, according to the conditions under which were carried out the experiments (pH and temperature), the non-dissociated ammonia concentration was below 56.5 mg NH3-N L-1 at pH 9.0 and 0.12 mg NH3-N L-1 at pH 6.5, i.e., far below the reported inhibitory concentrations for denitrification (Mosquera-Corral et al. 2001; Soto et al. 2007). Figure 2 shows the r/rmax ratio as a function of the pH in assays performed at an initial C/N of 10 and initial nitrate concentrations of 80 and 10 mg NO3--N L-1. Maximal nitrate consumption rates were obtained from the initial slope of the nitrate consumption vs. time plot at different pH values. As shown, the r/rmax ratio and, hence, the apparent maximum rate of nitrate consumption, reached a peak between pH 7.5 and 8.0 at initial nitrate concentrations of 10 mg NO3--N L-1 and 80 mg NO3--N L-1. At an initial nitrate concentration of 80 mg NO3--N L-1, the apparent maximum rate diminished 50% when the pH was decreased from 8.0 to 7.0 and when increased from 8.0 to 9.0. At an initial nitrate concentration of 10 mg NO3--N L-1, the apparent maximum rate diminished 50% when the pH increased from 8.0 to 9.0, but when pH decreased from 8.0 to 7.0, the apparent maximum rate diminished only 23%. This behaviour might be ascribed to different NO2--N and HNO2 concentrations. According to Soto et al. (2007), a 10% reduction in the rate of nitrate reduction at pH 7.5 is observed when the initial nitrate concentration is 74.24 mg NO3- L-1, i.e., the actual minimum inhibitory concentration of nitrous acid would be 0.004 mg HNO2-N L-1 (Soto et al. 2007). As shown in Table 1, the maximal HNO2 concentrations that might be reached at an initial nitrate concentration of 10 mg NO3--N L-1 and, assuming that all the initial nitrate is reduced to nitrite and that nitrite is not further transform to gaseous nitrogen, are lower than 0.004 mg HNO2 L-1 within the pH range of 6.5-9.0, whereas at 80 mg NO3--N L-1 the maximal HNO2 concentration is greater than 0.004 mg HNO2-N L-1 when the pH diminished to 7.5. Thus, assays performed at an initial nitrate concentration of 80 mg NO3--N L-1 and at a pH lower than 7.5 would reflect the combined HNO2 inhibitory effect and the pH inhibitory effect on the denitrification rate while assays performed at 80 mg NO3--N L-1 at pH values higher than 7.5 would only reflect the inhibitory effect of the pH on the denitrification rate. On the other hand, assays performed at an initial nitrate concentration of 10 mg NO3--N L-1 would reflect only the pH effect at the studied pH range. The optimum rate was attained at pH 7.5 to 8.0, and no differences in rates were observed between both initial nitrate concentrations. This pH range is in agreement with those reported by Glass et al. (1997), who reported an optimum pH for denitrification close to 8.0 in an active sludge reactor with a nitrate and nitrite concentrations of 1350 mg NO3--N L-1 and 15 - 2100 mg NO2--N L-1, respectively. Almeida et al. (1995) reported an optimum pH of 7.0 for denitrification carried out by a pure culture of Pseudomonas fluorescens at 28ºC. This optimum is lower than the one found in this work; the difference might be due to the different bacterial strains used, as in this work a mixed bacterial culture was used to perform the experiments. In fact, Kunak et al. (2004) utilizing a pure culture of Paracoccus denitrificans, found optimal pHs at 6.2 for nitrite reductase and 7.5 for nitric oxide reductase at 30-40ºC; these values are also different from those reported by Almeida et al. (1995). Modelling of the effect of the pH on the rate of nitrate reduction The effect of the pH on the apparent maximum rate of nitrate reduction at an initial nitrate concentration of 10 mg NO3--N L-1 was modelled by the Michaelis-type kinetics (equation 8). According to the Henderson-Hasselbach expression, the HNO2 concentration is negligible at the studied pH range; thus, at this nitrate concentration solely the pH affects the rate of denitrification. Figure 2 shows the fitting of the experimental data by the Michaelis model. The parameters pKs1 and pKs2 of the factor accounting for pH inhibition were calculated by fitting of the experimental data by equation (8). Fitting was carried out by the TableCurve 2D, program using the Levenberg-Marquardt method. The fitting of the experimental data gave the following parameter values: pKs1 = 6.27 ± 0.204; pKs2 = 9.04 ± 0.175; maximum specific rate at the optimum pH = 1.32 ± 0.24 [mg NO3--N (g VSS h)-1] and A = 1.01 ± 0.085. As shown in Figure 2, the function gives a good prediction of the r/rmax ratio at pH < 8.0, as the predicted values lie within the experimental error range. At pH values over 8 (8.5 and 9), the function was not able to predict values within the experimental error range. Effect of the pH on the apparent maximum specific rate of nitrite reduction Figure 3 shows the apparent maximum specific rates of nitrite reduction at initial nitrate concentrations of 80 mg NO3--N L-1 and 10 mg NO3--N L-1. Similarly as in nitrate reduction, the apparent maximum rate of nitrite consumption reached a peak between pH values of 7.5 and 8.0, at initial nitrate concentrations of 10 mg NO3--N L-1 and 80 mg NO3--N L-1. Hence, the optimum pH for nitrite reduction lies within these pH values. A decrease in pH from 8.0 to 6.5 or an increase in pH from 8.0 to 9.0 reduced by 40% the apparent nitrite consumption rate. This reduction can only be ascribed to a pH effect since a value of 0.05 mg HNO2-N L-1 was calculated as the substrate concentration at maximum specific growth rate, µmax from the kinetic parameters reported by Soto et al. (2007). Therefore, as the HNO2-N concentration never surpassed this value at pH values of 6.5-9.0 (Table 1), only limitation by substrate but not substrate inhibition could have existed. Modelling of the effect of the pH on the apparent maximum specific rate of nitrite reduction The separate effect of the pH on the maximum rate of nitrite reduction at an initial nitrate concentration of 10 mg NO3--N L-1 was modelled by the Michaelis function (equation 8). The parameters pKs1 and pKs2 of the factor accounting for pH inhibition were calculated by fitting of the experimental data by equation (8). Fitting was carried out by the TableCurve 2D, program using the Levenberg-Marquardt method. Figure 3 shows the fitting of the experimental data by this function for an initial nitrate concentration of 10 mg NO3--N L-1; as shown, this function fitted the experimental data in the whole pH range studied. The fitting of the experimental data gave the following parameter values: pKs1 = 6.41 ± 0.072, pKs2 = 8.93 ± 0.065, rmax = 1.07 ± 0.08 [mg NO2--N (g VSS h)-1] and A = 1.09 ± 0.024. Quantification of the inhibitory effect of the pH in reported kinetic modelsSeveral authors have modelled the nitrate consumption rate by the Monod equation. However, the kinetic parameters they reported were obtained with different carbon sources, at a different temperature and/or different pH than the conditions used in this work (mainly protein as carbon source, 37ºC and pH range 6.5-9.0). Wild et al. (1995) reported kinetic parameters at 20ºC with acetate as carbon source; moreover, Wild et al. (1995) performed their experiments at pH 7.0. On the other hand, Almeida et al. (1995) reported kinetic parameters for denitrification obtained at 28ºC, pH 7.0, with acetate as carbon source and using a pure culture of Pseudomonas fluorescens, while in this work denitrification was carried out by an anaerobic sludge adapted to nitrate. The kinetic parameters for the Monod model reported by Soto et al. (2007) were obtained at pH = 7.5 with the same inoculum and temperature used in this work although he used solely acetate as carbon source, thus, the experimental conditions used by Soto et al. (2007) were the closest to the ones used in this work. Therefore, the experimental nitrate and nitrite reduction kinetics at different pHs were compared to the Monod model with kinetic parameters reported by Soto et al. (2007). The constants for nitrate and nitrite reported by Almeida et al. (1995) and Soto et al. (2007) are shown in Table 2. The inclusion of the Michaelis function, which accounts for the effect of the pH, in the Monod and in the Haldane model gives the following equations: For nitrate reduction:
Similarly, for nitrite reduction:
Figure 4a and Figure 4b show the fittings by the Monod model and the pH-modified Monod model with the kinetic parameters reported by Soto et al. (2007) of the experimental data for nitrate consumption at pH 6.5 and 9.0, respectively, and at an initial nitrate concentration of 80 mg NO3--N L-1. Figure 4c and Figure 4d show the fitting by the Haldane model and the pH-modified Haldane model with the kinetic parameters reported by Soto et al. (2007) of the experimental data for nitrite accumulation at pH 6.5 and 9.0, respectively, and at an initial nitrate concentration of 10 mg NO3--N L-1. As shown, the Monod and Haldane equations with the reported parameters gave a poor fitting of the experimental data for nitrate reduction and nitrite accumulation rates, respectively. Table 3 show the deviations between the experimental data for nitrate and nitrite consumption and the fitting by the Monod and Haldane kinetic models with the empiric constants reported by Soto et al. (2007) before and after including the Michaelis function. As an average, deviation diminished from 33.8% to 10.5% for nitrate reduction and from 53% to 10.7% for nitrite reduction upon inclusion of the factor representing the pH inhibition in their kinetic expressions. No differences were observed between the predicted values obtained with the original kinetic expression and those predicted by the modified kinetic expression at pH 7.5, suggesting that the prediction of the nitrate and nitrite rates is not affected at this pH by the pH function. As shown, by introducing the Michaelis function, the fitting of the experimental data markedly improves, underlining the importance of including the pH effect on the denitrification kinetics. Modelling by the pH-modified Monod of the experimental data reported by Almeida et al. (1995) for nitrate and nitrite consumption at pH 7.0 validated the use of the Michaelis function to quantify the effect of the pH in the denitrification kinetic. The Michaelis parameters calculated from the data reported by Almeida et al. (1995) were pKS1 = 6.3; pKS2 = 7.8 and A = 1.4 for nitrate reduction and pKS1 = 6.4; pKS2 = 7.3 and A = 1.4 for nitrite reduction. The errors between the fitting by the Monod model and the pH-modified Monod model of the experimental data reported by Almeida et al. (1995) at pH 6.6, 7.4 and 7.8 were 2.77%, 4.15% and 2.74%, respectively, for nitrate reduction and 5.21%, 10.18% and 9.55%, respectively, for nitrite reduction. As shown, the errors are quite small except for the nitrite accumulation. The latter might be due to the fact that Almeida et al. (1995) used a very narrow pH range (6.6-7.8) and the calculated pKs1 and pKs2 values lie out of this pH range. This function allows modelling of the kinetics of denitrification by one equation that integrates kinetic variables and the pH effect. These results shows that the effect of the pH should be taken into account in the modelling, operation and design of bacterial processes.
The results of this work indicate that the inclusion of the pH effect through the Michaelis function in the kinetics of nitrate reduction and nitrite reduction clearly improves its predictive potential as it reduces the deviation between the predicted and experimental values from an average of 33.8% and 53.5% to 10.5% and 10.7% for nitrate reduction (Monod) and nitrite reduction (Haldane), respectively. Thus, not only the pH-dependent HNO2 effect should be considered in the modelling of the kinetics of nitrate and nitrite reduction but also the pH effect should be taken into account in these processes. ALMEIDA, J.S.; JULIO, S.M.; REIS, M.A.M. and LARRONDO, M.J.T. Nitrite inhibition of denitrification by Pseudomonas fluorescens. Biotechnology and Bioengineering, May 1995, vol. 46, no. 3, p. 194-201. [CrossRef] ANGELIDAKI, I.; ELLEGAARD, L. and AHRING, B. A mathematical model for dynamic simulation of anaerobic digestion of complex substrates: focusing on ammonia inhibition. Biotechnology and Bioengineering, June 1993, vol. 42, no. 3, p. 159-166. [CrossRef] APHA, AWWA, WPCF, Standard Methods for the Examination of Water and Wastewater. 20th ed. Amer. Public Health Assoc., Washington, D.C. 2002. ASPÉ, E.; MARTÍ, M.C.; JARA, A. and ROECKEL, M. Ammonia inhibition in the anaerobic treatment of fishery effluents. Water Environment Research, April 2001, vol. 73, no. 2, p. 154-164.[CrossRef] BATSTONE, D.; KELLER, J.; ANGELIDAKI, I.; KALYUZHNYI, S.; PAVLOSTATHIS, S.; ROZZI, A.; SANDERS, W.; SIEGRIST, H. and VAVILIN, V. Anaerobic Digestion Model Nº 1 (ADM1). London, IWA Task Group for Mathematical Modelling of Anaerobic Digestion Processes, Scientific and Technical Report Nº 13, 2002, 77 p. ISBN 1 900222 78 7. CAMPOS, E. and FLOTATS, X. Dynamic simulation of pH in anaerobic processes. Applied Biochemistry and Biotechnology, April-June 2003, vol. 109, no. 1-3, p. 63-76. [CrossRef] GLASS, C. and SILVERSTEIN, J. Denitrification kinetics of high nitrate concentration water: pH effect on inhibition and nitrite accumulation. Water Research, March 1998, vol. 32, no. 3, p. 831-839. [CrossRef] GLASS, C.; SILVERSTEIN, J. and OH, J. Inhibition of denitrification in activated sludge by nitrite. Water Environment Research, September-October 1997, vol. 69, no. 6, p. 1086-1093. [CrossRef] KUNAK, M.; KUCERA, I. and VAN SPANNING, R. Nitric oxide oscillations in Paracoccus denitrificans: the effects of environmental factors and of segregating nitrite reductase and nitric oxide reductase into separate cells. Archives of Biochemistry and Biophysics, September 2004, vol. 429, no. 2, p. 237-243. [CrossRef] MOSQUERA-CORRAL, A.; SANCHEZ, M.; CAMPOS, J.L.; MÉNDEZ, R. and LEMA, J.M. Simultaneous methanogenesis and denitrification of pretreated effluents from a fish canning industry. Water Research, February 2001, vol. 35, no. 2, p. 411-418. [CrossRef] RAMSAY, I.R. and PULLAMMANAPPALLIL, P.C. Full-scale application of a dynamic model for high-rate anaerobic wastewater treatment systems. Journal of Environmental Engineering-ASCE, July 2005, vol. 131, no. 7, p. 1030-1036. [CrossRef] RIVER, L.; ASPÉ, E.; ROECKEL, M. and MARTÍ, M.C. Evaluation of clean technology processes in the marine products processing industry.Journal of Chemical Technology and Biotechnology, November 1998, vol. 73, no. 3, p. 217-226.[CrossRef] SÁNCHEZ, O.; ASPÉ, E.; MARTI, M.C. and ROECKEL, M. Rate of ammonia oxidation in a synthetic saline wastewater by a nitrifying mixed-culture. Journal of Chemical Technology and Biotechnology, November 2005, vol. 80, no. 11, p. 1261-1267. [CrossRef] SEGEL, Irwin. Effects of pH and temperature. In: SEGEL, Irwined. Enzyme Kinetics. New York, USA, John Wiley & Sons, Inc., 1975, p. 884-941. SIEGRIST, H.; VOGT, D.; GARCIA-HERAS, J.L. and GUJER, W. Mathematical model for meso- and thermophilic anaerobic sewage sludge digestion. Environmental Science and Technology, March 2002, vol. 36, no. 5, p. 1113-1123. [CrossRef] SOTO, O.; ASPÉ, E. and ROECKEL, M. Kinetics of cross-inhibited denitrification of a high load wastewater. Enzyme and Microbial Technology, May 2007, vol. 40, no. 6, p. 1627-1634. [CrossRef] WILD, D.; VON SCHULTHESS, R. and GUJER, W. Structured modelling of denitrification intermediates. Water Science and Technology, 1995, vol. 31, no. 2, p. 45-54. [CrossRef] Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. |
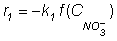 [2]
[2]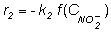 [3]
[3]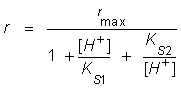 [6]
[6]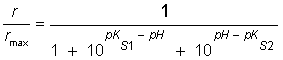 [7]
[7]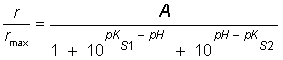 [8]
[8] [9]
[9] (pH-modified
Monod) [10]
(pH-modified
Monod) [10]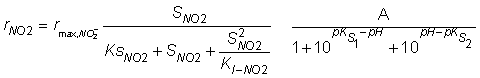 (pH-modified
Haldane) [11]
(pH-modified
Haldane) [11]