
|
Measuring β-Galactosidase activity at pH 6 with a differential pH sensor Cristian
Acevedo*# Matthias H. Stach Anette Amtmann Manuel E. Young Juan G. Reyes Holger Huebner Rainer Buchholz *Corresponding author Financial support: FONDEF Grant (DO2I1009), MECESUP Grant (UCV0206) and Doctoral Scholarship for Cristian Acevedo (CONICYT D-21050588). Present address: #Departamento de Ciencia y Tecnología de los Alimentos, Facultad Tecnológica, Universidad de Santiago de Chile. Avenida Ecuador 3769, Estación Central, Santiago, Chile. Keywords: β-Galactosidase, delta milli pH, differential pH.
The β-Galactosidase activity at pH 6 is used as a cellular marker to identify senescent cell cultures. The classic method to identify this enzymatic activity is using cytochemical staining with X-Gal after 16 hrs. In this work, a differential pH sensor was used to measure β-Galactosidase activity at pH 6. The measurement is easy and only takes 3 min.
Normal somatic cells invariably enter a state of irreversibly arrested growth and altered function after a finite number of divisions. This process, termed cellular senescence (Hayflick’s limit), is thought to be a tumor-suppressive mechanism and an underlying cause of aging. Normal cells express β-Galactosidase activity at pH 4, but senescent cells express β-Galactosidase activity at pH 6 (called senescence associated β-Galactosidase) (Dimri et al. 1995; Severino et al. 2000; Maier et al. 2007). The β-Galactosidase activity at pH 6 is used as a cellular marker to identify senescent cell cultures. The classic method to identify β-Gal/pH6 is using cytochemical staining with X-Gal (5-bromo-4-chloro-3-indolyl-β-D-galactoside) in cells fixed with glutaraldehyde/formaldehyde solution after 16-24 hrs (Dimri et al. 1995). But, other methods have been developed to measurement of β-Gal/pH6 activity (Bassaneze et al. 2008), such as the fluorescent assays using FDG (fluorescein di-β-D-galactopyranoside) (Yang and Hu, 2004) or MUG (4-methylumbelliferyl-β-D-galactopyranoside) in cell extract solutions (Gary and Kindell, 2005), because it is necessary to have quantitative and fast methodologies. On the other hand, the differential pH measurement technique is an analytical tool to detect and determine all sorts of metabolites in biological samples. Much faster to be performed and developed for a certain purpose than HPLC protocols, and much cheaper than most enzymatic test kits. Instead of determining the amount of metabolites, one can use this method to determine the amount of an enzyme of interest by adding the substrate in excess. The differential pH technique is based on the possibility to correlate the pH variation, induced by the change in the concentration of H+ or OH- by specific enzymatic reactions. The technique measures differences in the order of 10-3 units of pH using two micro glass electrodes of high sensitivity, reason why the study and definition of the system are essential for the standardization of new procedures. This technique has been used to measure substrates and enzymatic activities in biological systems with high precision and quickly (Compagnone et al. 1995; Luzzana and Giardino, 1999; Gast and Pingoud, 2001; Luzzana et al. 2001; Tagarelli et al. 2004). In this technical note, we developed a new technique for measuring β-Galactosidase activity at pH 6 using a differential pH sensor, which could be used in the future in cell culture technology and tissue engineering applications, in fast and quantitative way. The enzymatic system used consists of two reactions:
Since pKA values of ATP differ from
the pKA values of glucose-6P and ADP, the pH changes in the course of the
reaction (Luzzana et al. 2001). Our hypothesis is that the pH changes will be a
function of the amount of glucose, providing an indirect method for the
determination of the β-Galactosidase activity according to
Michaelis-Menten kinetics.
The Hexokinase solution was prepared at 0.2 U/µL and the β-Galactosidase standard solution at 0.1 U/µL. Both enzymes were purchased from Sigma-Aldrich. The reaction-buffer was prepared with citric-phosphate buffer (20 mM), ATP (15 mM), MgCl2 (2 mM) and NaCl (150 mM). The pH of reaction-buffer was checked and adjusted to 6.00 previously to use. The differential pH device used was the CL-10 plus (mode Init. String “CS4000”) (Eurochem, Italy). The kinetics in the differential pH electrodes was monitored for 200 sec at 37ºC. In the reaction chamber, 10 µL of lactose solution (substrate in excess: 100 mg/mL) with 10 µL of Hexokinase (10 µL x 0.2 U/µL = 2 U) were added. The measurement was started and immediately after the activation of the enzyme-pump, 10 µL of β-Galactosidase solution were injected into the reaction chamber (10 µL x 0.1 U/µL = 1 U). To measure the blank, lactose solution was replaced by pure water. The sample-kinetics (with substrate in excess) and blank-kinetics (without lactose) were measured per triplicate. The optimum pH of Hexokinase is 7.5 (Sols et al. 1958). In our experience, the differential pH sensor can detect the Hexokinase activity at this pH. However, to verify the response in an acid pH, the Hexokinase activity was checked adding 10 µL of glucose solution (0, 20 and 40 mM) in the reaction chamber using reaction-buffers adjusted to pH 4.00, 5.00 and 6.00. The measurement was started and 10 µL of Hexokinase solution were injected immediately after the activation of the enzyme-pump. In order to validate the differential pH technique in cells, the β-Galactosidase solution was replaced for a cell extract solution (Nasim and Trembath, 2005). Three cell extracts were obtained from three cultures incubated in T75 flask: cell line of 3T6 fibroblasts, human primary culture from early-passage (< 3) and long-term human primary culture. The extracts obtained (100 µL) were dissolved in 400 µL of reaction buffers (pH 6) and measure in the differential pH sensor. The early-passage culture showed a fast growth (doubling time < 30 hrs), indicating a non-senescence state. The long-term culture was considered those that showed arrest of cell growth. After roughly 50 population doublings the primary culture reached the arrest of the growth (evaluated as no change in cell counting after one week). As a criterion, it is accepted that human fibroblasts are senescent after 50 duplications. Our experience and previous work (Acevedo et al. 2009) indicate that this state is close to 90% of senescence evaluated by X-Gal staining. The system did not show activity at pH inferior to six. But, the system presented activity to pH 6. This is because the Hexokinase activity in that pH ranges - minor to six - is very low (Sols et al. 1958). The enzymatic activity of Hexokinase in the differential pH sensor was checked, confirming this one (Figure 1). Thus, it is not possible to determinate β-Galactosidase activity at other pH values using this enzymatic system, but those reactions are not important to determine β-Gal/pH6 activity. Nevertheless, this presents a limitation for the extension of methods at others pH values, for example, at physiological pH of lysosome (pH 4) (Dimri et al. 1995), when the normal β-Galactosidase is active. At pH 6, the delta-milli-pH kinetics (change of milli-pH by time) was always linear in the presence of lactose, and the slopes are increasing with the β-Galactosidase activity (Figure 2A). But, the system without substrate (lactose) also showed a weak signal, maybe by unspecific reactions. The unspecific reactions always were minor to the specific reactions. To remove the unspecific reaction of the assay, the blank (without substrate) was subtracted from sample-kinetics, showing a new purified kinetics with two sections (Figure 2B). The final section of the purified delta-milli-pH kinetics was linear and depends on the β-Galactosidase activity. The final sections (called slope in Figure 2B) are controlled by β-Galactosidase activity in the sample, because this enzyme produces glucose at constant velocity (v1), which is transformed by Hexokinase into glucose-6P (v2), changing the pH of the system:
In the first section of the kinetics, the delta-milli-pH slope is not linear because the glucose concentration increases with the time (v1 > v2). Then, the glucose kinetics is described by:
But, in the linear section (when the slope is constant), a steady-state is established because v1 = v2, and then, the glucose concentration is constant:
Using the results shown in Figure 2B and the relationship indicated in equation 6, an inverse plot of type Lineweaver-Burk was made showing a linear dependency of β-Gal/pH6 activity with the slope of the purified kinetics (sample-kinetics less blank-kinetics). The linear fit was significant (p < 0.05; ANOVA), with a Pearson correlation coefficient of 0.999 (Figure 2C). In order to validate this technique, the β-Galactosidase solutions were replaced for three cell extracts obtained from a cell line, early-passage primary culture and long-term primary culture. The differential pH responses of cell line and early-passage primary culture were not significant (p > 0.05). But, the long-term primary culture showed a significant change in differential pH response (p < 0.05). Those results are correct, because cell lines and young cells not expresses a senescence phenotype, but long-term cell cultures after 50 duplications have large amount of senescence cells (> 90%) expressing β-Galactosidase activity at pH 6 (Dimri et al. 1995). In conclusion, this simple method can be used to determine enzymatic activity of β-Galactosidase at pH 6 in diluted samples, such as samples of cell extract or other kind of solutions, in fast and quantitative way, because the measurement is easy and the time per measurement only takes 3 min. ACEVEDO, C.; BROWN, D.; YOUNG, M. and REYES, J. Senescent culture of human dermal-fibroblasts modify phenotype when immobilized in fibrin-polymer. Journal of Biomaterials Science - Polymer Edition. 2009. BASSANEZE, Vinícius; MIYAKAWA, Ayumi and KRIEGER, Jose. A quantitative chemiluminescent method for studying replicative and stress-induced premature senescence in cell cultures. Analytical Biochemistry, January 2008, vol. 372, no. 2, p. 198-203. [CrossRef] COMPAGNONE, D.; MOSCA, A. and PALLESCHI, G. Cyclic enzymatic determination of L-lactate by differential pH measurement. Analytical Letters, November 1995, vol. 28, no. 14, p. 2475-2490. DIMRI, G.; LEE, X.; BASILE, G.; ACOSTA, M.; SCOTT, G.; ROSKELLEY, C.; MEDRANOS, E.; LINSKENS, M.; RUBELJ, I.; PEREIRA-SMITH, O.; PEACOCKE, M. and CAMPISI, J. A biomarker that identifies senescent human cells in culture and aging skin in vivo. Proceedings of the National Academy of Sciences of the United States of America, September 1995, vol. 92, no. 20, p. 9363-9367. GARY, Ronald and KINDELL, Susan. Quantitative assay of senescence-associated β-galactosidase activity in mammalian cell extracts. Analytical Biochemistry, August 2005, vol. 343, no. 2, p. 329-334. [CrossRef] GAST, Frank and PINGOUD Alfred. Nucleic acid quantification by delta pH measurement. Journal of Biotechnology, June 2001, vol. 88, no. 2, p. 107-117. [CrossRef] LUZZANA, Massimo and GIARDINO, Rafaella. Urea determination in milk by a differential pH technique. Lait, April 1999, vol. 79, no. 2, p. 261-267. [CrossRef] LUZZANA, M.; AGNELLINI, D.; CREMONESI, P. and CARAMENTI, G. Enzymatic reactions for the determination of sugars in food samples using the differential pH technique. The Analyst, December 2001, vol. 126, no. 12, p. 2149-2152. [CrossRef] MAIER, Andrea; WESTENDORP, Rudi and VAN HEEMST, Diana. β-galactosidase activity as a biomarker of replicative senescence during the course of human fibroblast cultures. Annals of the New York Academy of Sciences, April 2007, vol. 1100, p. 323-332. [CrossRef] NASIM, M. and TREMBATH, R. A dual-light reporter system to determine the efficiency of protein-protein interactions in mammalian cells. Nucleic Acids Research, April 2005, vol. 33, no. 7, p. e66. [CrossRef] SEVERINO, J.; ALLEN, R.; BALIN, S.; BALIN, A. and CRISTOFALO, V. Is β-galactosidase staining a marker of senescence in vitro and in vivo? Experimental Cell Research, May 2000, vol. 257, no. 1, p. 162-171. [CrossRef] SOLS, Alberto; DE LA FUENTE, Gertrudis; VILLAR-PALASI, Carlos and ASENSIO, Carlos. Substrate specificity and some other properties of baker's yeast hexokinase. Biochimica et Biophysica Acta, October 1958, vol. 30, no. 1, p. 92-101. [CrossRef] TAGARELLI, Antonio; PIRO, Anna; TAGARELLI, Giuseppe; BASTONE, Loredana; PALEARI, Renata and MOSCA, Andrea. G6PD/PK ratio: a reliable parameter to identify glucose-6-phosphate dehydrogenase deficiency associated with microcytic anemia in heterozygous subjects. Clinical Biochemistry, October 2004, vol. 37, no. 10, p. 863-866. [CrossRef] YANG, N. and HU, M. A fluorimetric method using fluorescein di-β-D-galactopyranoside for quantifying the senescence-associated β-galactosidase activity in human foreskin fibroblast Hs68 cells. Analytical Biochemistry, February 2004, vol. 325, no. 2, p. 337-343. [CrossRef] Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. |
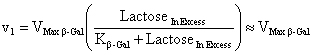 [Eq. 3]
[Eq. 3]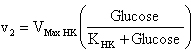 [Eq. 4]
[Eq. 4]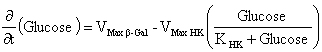 [Eq. 5]
[Eq. 5]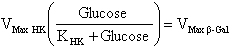 [Eq. 6]
[Eq. 6]