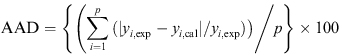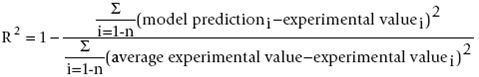|
Thermostable lipase from a newly isolated Staphylococcus xylosus strain; process optimization and characterization using RSM and ANN Anahita Khoramnia Oi Ming Lai* Afshin Ebrahimpour Carynn Josue Tanduba Tan Siow Voon Suriati Mukhlis *Corresponding author Financial support: University Putra Malaysia. Keywords: artificial neural network, characterization, lipase, optimization, response surface methodology, Staphylococcus xylosus.
Normal feed forward back-propagation artificial neural network (ANN) and cubic backward elimination response surface methodology (RSM) were used to build a predictive model of the combined effects and optimization of culture parameters for the lipase production of a newly isolated Staphylococcus xylosus. The results demonstrated a high predictive accuracy of artificial neural network compared to response surface methodology. The optimum operating condition obtained from the ANN model was found to be at 30ºC incubation temperature, pH 7.5, 60 hrs incubation period, 1.8% inoculum size and 60 rpm agitation. The lipase production increased 3.5 fold for optimal medium. The produced enzyme was characterized biochemically and this is the first report about a mesophilic staphylococci bacterium with a high thermostable lipase which is able to retain 50% of its activity at 70ºC after 90 min and at 60ºC after 120 min. This lipase is also acidic and alkaline resistant which remains active after 24 hrs in a broad range of pH (4-11).
Lipolytic enzymes catalyse hydrolysis and synthesis reactions, either in long chain triacylglycerides (lipases) or in short chain fatty acids (esterases) (Arpigny and Jaeger, 1999). Nowadays, thermozymes are gaining wide industrial and biotechnological interest due to the fact that they are better suited for harsh industrial processes (Jaeger and Eggert, 2002; Sharma et al. 2002). Recently thermostable lipases from various microorganisms have been purified and characterized such as Bacillus spp. and Geotrichum candidum Y05 (Berger et al. 1995; Fuciños et al. 2008). Microbial thermostable lipases are of considerable commercial interest for biotechnological applications as they can be produced at low cost (Schmidt-Dannert et al. 1996). In the present work, we isolated a mesophilic thermostable lipase producing bacterium from food oil waste in Malaysia that has been identified as Staphylococcus xylosus. Staphylococci lipases play an important role in the development of aroma of fermented products (Talon and Montel, 1997). S. xylosus, is commonly used as lipolytic starter cultures for fermented meat products such as sausages and ham (Jessen, 1995). Staphylococci lipases are mostly extracellular and like other lipases, their production influenced by several nutritional and physical factors, namely the carbon source, nitrogen source, temperature, pH, dissolved oxygen concentration and presence of inducers (Ebrahimpour et al. 2008), here we focused on the physical factors. Fermentation processes are used to produce various substances in the pharmaceutical, chemical and food industries. The engineering of culture conditions is also an effective mode to achieve enzyme preparations that are enriched in selected isoenzymes which are effective for particular biotechnological applications (Hsu et al. 2008). Modeling and optimization are the most important stages in biological processes to increase the yield of the process and to improve a system without increasing the cost (Baş and Boyaci, 2007). Various process optimization strategies were developed by scientists and the most frequently used optimization strategy is “one-at-a-time” strategy (Dutta et al. 2004). This classical method is not only time consuming, but also does not depict the combined interactions between physio-chemical parameters and may lead to misinterpretation of results (Baş and Boyaci, 2007; Sim and Kamaruddin, 2008). To overcome these problems, modern optimization systems are employed. Response surface methodology (RSM) is the most widely used models which includes factorial design and regression analysis. It helps in building models and in evaluating the effective factors to study interaction, and select optimum conditions of variables for a desired response (Liu et al. 1999; Vohra and Satyanarayana, 2002). Recently, a number of statistical experimental designs with RSM have been employed for optimizing conditions which the most widely used simulating models are second order polynomials (Lou and Nakai, 2001; He and Xiao, 2004). In the last decade, artificial neural networks (ANNs) have emerged as an attractive tool for developing non-linear models especially in situations where in the development of conventional empirical models becomes impractical (Patnaik, 2006). Lou and Nakai (2001) used an artificial neural network (ANN) method, computer model system, which match the functionality of the brain in a fundamental manner, represents the nonlinearities in a much better way. Liu et al. (2009) demonstrated that the ANN has good predictability and accuracy in optimizing the multi-factor, non-linear, and time-variant bioprocess. Although different reports showed lipase production optimization by various bacteria, in the case of optimization of Staphylococcal lipase production, ours is the first, consequently optimization using ANN is also the first. There are some reports about enzyme production optimization using ANN (Dutta et al. 2004; Ebrahimpour et al. 2008). In the present study, experiments were designed and optimized to allow for a cost-effective fermentation medium for lipase production from a newly isolated S. xylosus. For this purpose, the selected factors were optimized by RSM and ANN as comparison using central composite rotatable design (CCRD) and subsequently the produced enzyme was characterized biochemically. Screening of lipolytic micro-organisms The screening was realized on a nutrient agar medium containing 1% olive oil and 1% Rhodamin B. The culture plates were incubated at 30ºC, and colonies giving rise to widespread clearing around them under UV were considered as lipase producers. The bacterial strain used in this study was isolated from oily food waste in Serdang, Selangor, Malaysia and identified as Staphylococcus xylosus via BIOLOG system as well as biochemical properties and the morphological aspect. This new strain was preserved in sterile 16% (v/v) glycerol in Tryptic Soy Broth (TSB) at -80ºC. Composition of lipase production medium The composition of the selected lipase production medium was as follows (% w/v): peptone (5), yeast extract (1), NaCl (0.05), CaCl2 (0.05), lactose (1) and olive oil (1 v/v). The medium was sterilized for 15 min at 121ºC. The physical factors were fixed according to experimental design. After lipase production, the cell-free supernatant was obtained by centrifugation at 12,000 g, 4ºC and 5 min prior to lipase assay. Determination of liberated free fatty acid was measured by colorimetric assay (Kwon and Rhee, 1986) using olive oil as substrate. The enzymatic reaction was performed in a water bath shaker for 30 min at 30ºC under 250 rpm agitation. One unit of lipase activity was defined as 1.0 Uml-1 of free fatty acid liberated min-1 and reported as Uml-1. A five-level-five-variable CCRD was employed in this study, requiring 26 experiments (Cohran and Cox, 2002). The variables (physical factors) and their levels selected for the lipase production optimization were: T, incubation temperature (27 - 45ºC); pH, initial pH (6 - 9); IS, inoculum size (1 - 5%); Agt, agitation rate (0 - 200 rpm) and t, incubation period (24 - 96 hrs). Response surface methodology analysis In order to find the best polynomial equation of production media physical parameters, CCRD was employed for model fitting in RSM. The experimental data requiring 26 points according to CCRD (Table 1) was analyzed using Design Expert version 6.0.6 (Stat Ease Inc. Minneapolis, USA) and then interpreted and all tests were performed in triplicate. Three main analytical steps: analysis of variance (ANOVA), regression analysis and plotting of response surface were carried out to obtain the optimum condition for the lipase production. In order to test the model, the actual values obtained experimentally were compared to the RSM model predicted values. Finally, the predicted optimum values were tested experimentally to validate the reliability of the model. Artificial neural network analysis A commercial ANN software, NeuralPower version 2.5 (CPC-X Software) was used for the lipase optimization in this study. Same set of data (CCRD) was used for ANN but, instead of 5 replicates in centre point, average of center points was used (22 run instead of 26 run). It is well established that unlike RSM, the replicates at center point do not improve the prediction capability of the network in ANN modeling, because of the similar inputs (Baş and Boyaci, 2007). Thus the experimental data requiring 22 points (Table 1) was divided into two sets: training and testing sets. Hence, 19 out of 22 run of the CCRD experimental data was used as training sets. In order to predict the lipase activity, multilayer normal feed forward and multilayer full feed forward neural networks were employed separately. Different learning algorithms (incremental back propagation, IBP; batch back propagation, BBP; quickprob, QP; genetic algorithm, GA; and Levenberg-Marquardt algorithm, LM) were used for training the networks. Each ANN was trained until the network root of mean square error (RMSE) was lower than 0.001, average correlation coefficient (R) and average determination coefficient (DC) were equal to 1. Other ANN parameters were chosen as the default values of the software (Ebrahimpour et al. 2008). Three remaining points (out of 22) were used to test the obtained network. Finally, experimental value of the predicted optimal condition (Table 2) was used as validating test. All tests were performed in triplicate. Validation of the optimized condition In order to test the reliability of the estimation capabilities of the employed techniques, the predicted responses obtained from RSM and ANNs were compared with the experimental values (Table 3). The coefficient of determination (R2) and absolute average deviation (AAD) were determined and these values were used together to evaluate and compare ANN model with RSM. The AAD and R2 are calculated by Equation 1 and Equation 2, respectively. [Equation 1] [Equation 2] where yi,exp and yi,cal are the experimental and calculated responses, respectively, and p is the number of the experimental run. n is the number of experimental data. R2 is a measure of the amount of the reduction in the variability of response obtained by using the repressor variables in the model. Since R2 alone is not a measure of the model’s accuracy, it is necessary to use AAD analysis, which is a direct method for describing the deviations. Evaluation of R2 and AAD values together would be better to check the accuracy of the model. R2 must be close to 1.0 and the AAD between the predicted and observed data must be as small as possible (Baş and Boyaci, 2007, Ebrahimpour et al. 2008). The acceptable values of R2 and AAD values mean that the model equation defines the true behavior of the system and it can be used for interpolation in the experimental domain (Baş and Boyaci, 2007). Effect of pH and temperature on S. xylosus lipase activity and stability In order to find the optimum pH, S. xylosus lipase activity was tested at different pH (6-9) at standard screening temperature (37ºC), under 250 rpm agitation for 30 min. The optimum temperature for lipase activity was determined by carrying out the enzyme assay at different temperatures (25-70ºC) at optimum pH (8) for 30 min, under 250 rpm agitation. The pH stability was determined by incubating the enzyme at different pH (4-11) for 24 hrs at optimum temperature (40ºC) following by lipase assay at optimum condition. The thermostability was determined by incubating the enzyme solution at different temperatures (50-70ºC) at optimum pH (8) for 120 min following by lipase assay at optimum condition. The residual activity was determined using standard assay method (Kwon and Rhee, 1986) in triplicates. The central composite rotatable design (CCRD) along with the observed responses shown in Table 1, which are the lipase concentration produced by S. xylosus. The independent variables were: incubation temperature (T), initial pH (pH), inoculum size (IS), agitation rate (Agt) and incubation period (t). Response surface methodology analysis of lipase production of S. xylosus Five repeated experiments at the centre (0, 0, 0) of the design (run number 22-26 in Table 1) were performed to allow the estimation of the pure error. All experiments were carried out in a randomized order to minimize the effect of unexpected variability in the observed response due to extraneous factors (Shao et al. 2007). The data were fitted to various models (linear, two factorial, quadratic, cubic, modified) and according to their subsequent ANOVA the best fitted model was multiple linear regressions with backward elimination. Through this multiple linear regressions backward elimination, insignificant factors and interactions were removed from the model step by step and it was refitted. In each subsequent step, the least significant variable in the model was removed until all remaining variables had individual P-values smaller than 0.05 (except the main factors due to hierarchy) (Cheong et al. 2007). The final model was a modified cubic model (Equation 3) and its subsequent ANOVA (Table 4) showed a well-fitted model for optimization of lipase production. Lipase activity (Uml-1) = -4.88 + 0.11 T + 0.6 pH - 0.14 IS + 0.025 t + 3.85 E-3 Agt -2.33 E-4 T2 - 0.021 pH2 - 0.015 IS2 - 6.59 E-5 t2 - 4.94 E-6 Agt2 -8.32 E-3 t.pH - 3.59 E-4 T.t -1.13 E-4 T.Agt + 0.017 pH.IS - 9.02 E-4 pH.t + 6.97 E-4 IS.t + 3.7 E-4 IS.Agt where T is temperature, IS is inoculum size, Agt is agitation rate, t is incubation period and pH is initial medium pH. With very small "model P-value" (0.0009) and large "lack of fit P-value" (0.2942) from the analysis of ANOVA and a suitable coefficient of determination (R2 = 0.9589) and adjusted coefficient of determination (R2 adjusted = 0.8714), the modified cubic polynomial model was well fitted and statistically applicable to represent the actual relationship between the response and the significant variables (Table 4). In order to evaluate the fitness of this cubic model, comparison of predicted and experimental data sets were made by calculation of R2 and AAD and the obtained values were 0.9589 and 10.512%, respectively (Table 3). Artificial neural network analysis and modeling The best ANN chosen in the present work was a multilayer normal feed forward incremental back propagation network with optimized values of network learning rate and momentum of 0.15 and 0.8, respectively. The best topology was Gaussian transfer function consisted of a 5-15-1 (inputs- hidden layers -output) topology. The learning was accomplished in RMSE < 0.001, R = 1 and DC = 1. In order to evaluate the fitness of the ANN model employed, comparison of predicted and experimental values for training, testing and validating data sets were made by calculation of R2 and AAD. In the case of training data set, the coefficient of determination (R2) was 0.997206 and the related absolute average deviation (AAD) was 2.9809%, whereas for the testing data set (italic digits), these values were 0.996755and 4.7204%, respectively (Table 3). Comparison of RSM and ANN predicted values According to the obtained statistical parameters of R2 and AAD for training and testing data sets of ANN and whole data set of RSM (Table 3), we can conclude that ANN is capable in data fitting, prediction and generalization of both known (training data) and unknown (testing) data sets when compared to RSM. Also in the case of validating of the models, we obtained the similar result. As shown in Table 2, optimum points of RSM and ANN which were selected as the highest amount of the predicted lipase production among a set of solutions (data not shown) are used as validating test and the experimental values are the average of triplicates. Though both models preformed well and offered stable responses, again the ANN based approach was better in estimation capabilities and optimum point determination in comparison to the RSM. Also it is shown by Dutta et al. (2004) and Ebrahimpour et al. (2008) that ANNs are superior and more accurate modeling techniques when compared to RSM. Main effects and interactions between parameters Three dimensional response surface curves were plotted to study the optimum level as well as its interaction effect with other parameters of each variable on lipase production. Each response surface plot represents the effect of two independent variables at a constant level of the other variables. Many parameters can influence the lipase production. The most important parameters which had significant effects on lipase production of S. xylosus were incubation temperature (T), inoculum size (IS) and agitation (Agt) according to the ANOVA analysis (Table 4) and importance of parameters graph (Figure 1). ANOVA analysis show that although pH and incubation period were not significant parameters (P value > 0.05), they had significant interactions with other parameters, hence it has been used to develop the model. Figure 2a shows that lipase activity effectively increased with a decrease in inoculum size but temperature did not show significant effect on lipase production. On the other hand, ANOVA analysis and Figure 1 revealed, that temperature is one of the most important parameters for lipase production. As a conclusion, though both inoculum size and temperature parameters are significant, yet their interaction is not a significant parameter for lipase production. Hence modification of model via the removal of this interaction using backward elimination strategy improved the model (Equation 3). Figure 2b represents the three dimensional plot showing the interaction of temperature and pH on lipase production. Maximum lipase activity was obtained at 30ºC and pH 7.5. Further increase or decrease in the temperature and pH, led to the decrease in the enzyme production. Generally, the enzymatic synthesis can be greatly associated with cell growth (Fuciños et al. 2008; Deive et al. 2009). It was shown by Mauriello et al. (2004) that the growth temperature of Staphylococcus sp. was between 30 and 37ºC. Thus, the highest value of enzyme production, maximum at 30ºC was also attained in the same range of temperature values where the microorganism showed optimal growth. According to the plot, a neutral pH (7.5) caused maximum production and it was concluded that the microorganism needs neutral or alkaline pH values for production. These results could be expected, considering that these microorganisms were isolated from oil wastes of food and natural habitats where the pH is neutral or alkaline (Kristjánsson and Hreggvidsson, 1995). Moreover most bacteria prefer neutral initial pH for the best growth and lipase production (Baird-Parker, 1990). Figure 2c shows the initial pH and inoculum size interaction effects on the lipase activity. As shown in this figure and also according to the equation, the optimum amount of inoculum size for lipase activity is 1.8%. In order to enhance the growth of bacteria and the lipase production, inoculum size must be suitable and therefore, the nutrient and oxygen levels are sufficient (Ebrahimpour et al. 2008). Figures 2d and 2e show the inoculum size-agitation rate and time-temperature interactions, respectively. These plots reveal that the optimal lipase activity is at 60 hrs of incubation time and 60 rpm of agitation. Any decreases or increases in these amounts decrease the lipase activity. Lipases are produced throughout bacterial growth, with optimum production at late exponential growth phase (Gupta et al. 2004). Therefore, depending on environmental conditions and characteristics of the microorganism itself, the optimum incubation time is based on duration of log phase where limitation of growth elements occur and this can be an inducer for the production of some enzymes (Ebrahimpour et al. 2008). Generally, suitable agitation lead to increase of nutrient uptake and dissolved oxygen in the media (Kumar and Takagi, 1999). However, it has been demonstrated that moderate aeration (low shaking) produced much more lipase than high aeration (high shaking) (Rahman et al. 2006). Validation of the optimized condition The optimal conditions for lipase production were predicted as presented in Table 2 along with their predicted and actual values for both RSM and ANN. Among the various optimum conditions, the highest lipase activity (0.3594 Uml-1; 3.5-fold increase) was obtained using ANN at the following condition; growth temperature (30ºC), inoculum size (1.8%), agitation rate (60 rpm), incubation period (60 hrs) and initial pH (7.5). Effects of pH on lipase activity and stability The importance of acid and alkaline conditions on thermostable lipases has been studied for different applications. The activity of S. xylosus was investigated at different pH using olive oil as substrate. Unlike previously described staphylococcal lipases (Rosenstein and Götz, 2000), our results showed that S. xylosus lipase remained active at a pH range of 6-10. Under same experimental conditions, the S. xylosus lipase activity was maximal at pH 8 (Sayari et al. 2001; Mosbah et al. 2005) (Figure 3). A great deal of research is currently going into developing lipases which will work under alkaline conditions as fat stain removers. In pH stability study, the lipase was stable at a broad range of pH values between pH 4 and 11 after 24 hrs incubation. Under same experimental conditions, some other Staphylococcal lipases were found to be stable between pH 4 and 9 (Sayari et al. 2001; Mosbah et al. 2005). Lipases which are active and stable in acid and alkaline media are very attractive as these enzymes have a great potential for different industrial applications such as leather manufacture, detergent formulation, pulp and paper manufacture, and dairy industry (Jaeger and Eggert, 2002; Hasan et al. 2006). Effects of temperature on lipase activity and stability The S. xylosus lipase showed the highest activity at 40ºC (Figure 4). It was stable at 70ºC after 90 min and at 60ºC after 120 min (Figure 5). Sayari et al. (2001) reported that a Staphylococcal lipase was inactivated after a few minutes when incubated at 60°C while, Mosbah et al. (2005) and Horchani et al. (2009) reported the optimum activity at 45ºC and 55ºC and stability after 15 min and 60 min incubation at 60ºC respectively. Although our S. xylosus lipase showed the optimum activity at lower temperature (40ºC) but, the stability was much better. Effects of calcium on lipase activity Metal cations, particularly Ca2+, play important roles in influencing the structure and function of enzymes, and calcium-stimulated lipases have been reported (El Khattabi et al. 2003). It has been demonstrated that the activity of Staphylococcal lipases may depend on the presence of Ca2+ ions (Rosenstein and Götz, 2000). The effect of various Ca2+ concentrations on the rate of hydrolysis of emulsified olive oil by this lipase was studied. Our results showed that lipase activity could be detected in the absence of Ca2+ however, the enzymatic activity of this staphylococcal lipase was stimulated by Ca2+ (Figure 6). The lipase activity of S. xylosus increased maximum about 3 times at the Ca2+ concentration of 10 mM however, Mosbah et al. (2005) reported 1.9 times increase with 2 mM Ca2+ concentration. It has been reported that the lipases from Staphylococcushyicus (Rosenstein and Götz, 2000; Tiesinga et al. 2007) contain a Ca2+-binding site which is formed by two conserved aspartic acid residues near the active-site, and that binding of the Ca2+ ion to this site dramatically enhanced the activities of these enzymes. Our study attempted to show that Ca2+ does not seem to be necessary to detect the S. xylosus lipase activity but, it requires the presence of Ca2+ to enhance the hydrolysis of triacylglycerols. This study compared the performance of the central composite design of RSM and ANN in the estimation of operating parameters of lipase production from a newly isolated S. xylosus. Normal feed forward back-propagation ANN has been proven to be significantly more accurate in modeling, estimation and optimum point prediction than RSM. The final composition of the optimized culture conditions using ANN was as follows: 30ºC incubation temperature, initial pH 7.5, incubation period 60 hrs, inoculum size 1.8% and agitation 60 rpm which resulted in an overall 3.5-fold increase compared with that using the original conditions. The results suggest the feasibility of the application of ANN as the strategy to increase extracellular lipolytic enzyme production and decrease the cost. The knowledge and information obtained here may be also helpful to the other industrial bioprocesses to improve productivity. The produced enzyme was characterized biochemically and it showed better temperature and pH stability compared to previous studies on staphylococcal lipases. Also the presence of Ca2+ was not necessary for this lipase detection however it improved the hydrolysis of triacylglycerols. Thus, such a S. xylosus strain may have highly potential application for industrial production of a novel thermostable lipase. In addition this lipase can be useful to produce new products as well as other benefits of microbial thermostable lipases however, additional studies are necessary to evaluate its performance directly in different reactions catalysis. ARPIGNY, Jean Louis and JAEGER, Karl-Erich. Bacterial lipolytic enzymes: classification and properties. Biochemical Journal, July 1999, vol. 343, no. 1, p. 177-183. [CrossRef] BAIRD-PARKER, A.C. The staphylococci: an introduction. Journal of Applied Microbiology, January 1990, vol. 69, no. S19, p. 1S-8S. [CrossRef] BAŞ, Deniz and BOYACI, Ismail H. Modeling and optimization I: Usability of response surface methodology. Journal of Food Engineering, February 2007, vol. 78, no. 3, p. 836-845. [CrossRef] BERGER, J.L.; LEE, B.H. and LACROIX, C. Identification of new enzyme activities of several strains of Thermus species. Applied Microbiology and Biotechnology, December 1995, vol. 44, no. 1-2, p. 81-87. [CrossRef] CHEONG, Ling-Zhi; TAN, Chin-Ping; LONG, Kamariah; YUSOFF, Mohd Suria Affandi; ARIFIN, Norlelawati; LO, Seong-Koon and LAI, Oi-Ming. Production of a diacylglycerol-enriched palm olein using lipase-catalyzed partial hydrolysis: Optimization using response surface methodology. Food Chemistry, 2007, vol. 105, no. 4, p. 1614-1622. [CrossRef] COHRAN, W.G. and COX, G.M. Experimental Design. New York, Wiley, 2002, 613 p. ISBN 0-471-54567-8. DEIVE, Francisco J.; CARVALHO, Elisabete; PASTRANA, Lorenzo; RUA, Maria L.; LONGO, Maria A. and SANROMAN, M. Angeles. Strategies for improving extracellular lipolytic enzyme production by Thermus thermophilus HB27. Bioresource Technology, July 2009, vol. 100, no. 14, p. 3630-3637. [CrossRef] DUTTA, Jayati Ray; DUTTA, Pranab Kumar and BANERJEE, Rintu. Optimization of culture parameters for extracellular protease production from a newly isolated Pseudomonas sp. using response surface and artificial neural network models. Process Biochemistry, October 2004, vol. 39, no. 12, p. 2193-2198. [CrossRef] EBRAHIMPOUR, Afshin; RAHMAN, Raja Noor Zaliha Raja Abd; HOOI EAN CH'NG, Diana; BASRI, Mahiran and SALLEH, Abu Bakar. A modeling study by response surface methodology and artificial neural network on culture parameters optimization for thermostable lipase production from a newly isolated thermophilic Geobacillus sp. strain ARM. BMC Biotechnology, December 2008, vol. 8, no. 1, p. 96. [CrossRef] EL KHATTABI, Mohamed; VAN GELDR, Patrick; BITTER, Wilbert and TOMMASSEN, Jan. Role of the calcium ion and the disulfide bond in the Burkholderia glumae lipase. Journal of Molecular Catalysis B: Enzymatic, July 2003, vol. 22, no. 5-6, p. 329-338. [CrossRef] FUCIÑOS, Pablo; RUA, M. Luisa; LONGO, Maria A.; SANROMAN, M. Angeles and PASTRANA, Lorenzo. Thermal spring water enhances lipolytic activity in Thermus thermophilus HB27. Process Biochemistry, December 2008, vol. 43, no. 12, p. 1383-1390. [CrossRef] GUPTA, R.; GUPTA, N. and RATHI, P. Bacterial lipases: an overview of production, purification and biochemical properties. Applied Microbiology and Biotechnology, June 2004, vol. 64, no. 6, p. 763-781. [CrossRef] HASAN, Fariha; SHAH, Aamer Ali and HAMEED, Abdul. Industrial applications of microbial lipases. Enzyme and Microbial Technology, June 2006, vol. 39, no. 2, p. 235-251. [CrossRef] HE, Zuo and XIAO, Yuexiang. Regression-based artificial neural network methodology in response surface methodology. Transactions of Tianjin University, 2004, vol. 10, no. 2, p. 153-157. HORCHANI, Habib; MOSBAH, Habib; BEN SALEM, Nadia; GARGOURI, Youssef and SAYARI, Adel. Biochemical and molecular characterisation of a thermoactive, alkaline and detergent-stable lipase from a newly isolated Staphylococcus aureus strain. Journal of Molecular Catalysis B: Enzymatic, April 2009, vol. 56, no. 4, p. 237-245. [CrossRef] HSU, Ken-Hao; LEE, Guan-Chin and SHAW, Jei-Fu. Promoter analysis and differential expression of the Candida rugosa lipase gene family in response to culture conditions. Journal of Agricultural and Food Chemistry, March 2008, vol. 56, no. 6, p. 1992-1998. [CrossRef] JAEGER, Karl-Erich and EGGERT, Thorsten. Lipases for biotechnology. Current Opinion Biotechnology, August 2002, vol. 13, no. 4, p. 390-397. [CrossRef] JESSEN, B. Fermented Meats. In: CAMPBELL-PLATT, G. and COOK, P.E. Eds. Starter cultures for meat fermentation. Glasgow, Blakie Academic & Professional, 1995. p. 130-154. KRISTJÁNSSON, J.K. and HREGGVIDSSON, G.O. Ecology and habitats of extremophiles. World Journal of Microbiology and Biotechnology, January 1995, vol. 11, no. 1, p. 17-25. [CrossRef] KUMAR, C. Ganesh and TAKAGI, Hiroshi. Microbial alkaline proteases: from a bioindustrial viewpoint. Biotechnology Advances, December 1999, vol. 17, no. 7, p. 561-594. [CrossRef] KWON, Dae Y. and RHEE, Joon S. A simple and rapid colorimetric method for determination of free fatty acids for lipase assay. Journal of the American Oil Chemists’ Society, January 1986, vol. 63, no. 1, p. 89-92. [CrossRef] LIU, Chi-Hsien; HWANG, Chin-Fa and LIAO, Chii-Cherng. Medium optimization for glutathione production by Saccharomyces cerevisiae. Process Biochemistry, January 1999, vol. 34, no. 1, p. 17-23. [CrossRef] LIU, Long; SUN, Jun; ZHANG, Dongxu; DU, Guocheng; CHEN, Jian and XU, Wenbo. Culture conditions optimization of hyaluronic acid production by Streptococcus zooepidemicus based on radial basis function neural network and quantum-behaved particle swarm optimization algorithm. Enzyme and Microbial Technology, January 2009, vol. 44, no. 1, p. 24-32. [CrossRef] LOU, Wengao and NAKAI, Shuryo. Application of artificial neural networks for predicting the thermal inactivation of bacteria: a combined effect of temperature, pH and water activity. Food Research International, 2001, vol. 34, no. 7, p. 573-579. [CrossRef] MAURIELLO, G.; CASABURI, A.; BLAIOTTA, G. and VILLANI, F. Isolation and technological properties of coagulase negative staphylococci from fermented sausages of Southern Italy. Meat Science, May 2004, vol. 67, no. 1, p. 149-158. [CrossRef] MOSBAH, Habib; SAYARI, Adel; MEJDOUB, Hafedh; DHOUIB, H. and GARGOURI, Youssef. Biochemical and molecular characterization of Staphylococcus xylosus lipase. Biochimica et Biophysica Acta (BBA)-General Subjects, May 2005, vol. 1723, no. 1-3, p. 282-291. [CrossRef] PATNAIK, P.R. Synthesizing cellular intelligence and artificial intelligence for bioprocesses. Biotechnology Advances, March-April 2006, vol. 24, no. 2, p. 129-133. [CrossRef] RAHMAN, Raja Noor Zaliha Raja Abdul; BAHARUM, Sarul Nataqain; SALLEH, Abu Bakar and BASRI, Mahiran. S5 Lipase: An organic solvent tolerant enzyme. The Journal of Microbiology, 2006, vol. 44, no. 6, p. 583-590. ROSENSTEIN, Ralf and GÖTZ, Frederich. Staphylococcal lipases: Biochemical and molecular characterization. Biochimie, November 2000, vol. 82, no. 11, p. 1005-1014. [CrossRef] SAYARI, Adel; AGREBI, Nouredinne; JAOUA, Samir and GARGOURI, Youseff. Biochemical and molecular characterization of Staphylococcus simulans lipase. Biochimie, September 2001, vol. 83, no. 9, p. 863-871. [CrossRef] SCHMIDT-DANNERT, Claudia; RUA, Maria Luisa; ATOMI, Haruyuki and SCHMID, Rolf D. Thermoalkalophilic lipase of Bacillus thermocatenulatus. I. molecular cloning, nucleotide sequence, purification and some properties. Biochimica et Biophysica Acta (BBA)-Lipids and Lipid metabolism, May 1996, vol. 1301, no. 1-2, p. 105-114. [CrossRef] SHAO, P.; JIANG, S.T. and YING, Y.J. Optimization of molecular distillation for recovery of tocopherol from rapeseed oil deodorizer distillate using response surface and artificial neural network models. Food and Bioproducts Processing, June 2007, vol. 85, no. 2, p. 85-92. [CrossRef] SHARMA, R.; SONI, S.K.; VOHRA, R.M.; JOLLY, R.S.; GUPTA, L.K. and GUPTA, J.K. Production of extracellular alkaline lipase from a Bacillus sp. RSJ1 and its application in ester hydrolysis. Indian Journal Microbiology, 2002, vol. 42, p. 49-54. SIM, Jia Huey and KAMARUDDIN, Azlina Harun. Optimization of acetic acid production from synthesis gas by chemolithotrophic bacterium-Clostridium aceticum using statistical approach. Bioresource Technology, May 2008, vol. 99, no. 8, p. 2724-2735. [CrossRef] TALON, R. and MONTEL, M.C. Hydrolysis of esters by staphylococci. International Journal of Food Microbiology, May 1997, vol. 36, no. 2-3, p. 207-214. [CrossRef] TIESINGA, Jan J.W.; POUDEROYEN, Van Gertie; NARDINI, Marco; RANSAC, Stéphane and DIJKSTRA, Bauke W. Structural basis of phospholipase activity of Staphylococcus hyicus lipase. Journal of Molecular Biology, August 2007, vol. 371, no. 2, p. 447-456. [CrossRef] VOHRA, A. and SATYANARAYANA, T. Statistical optimization of the medium components by response surface methodology to enhance phytase production, Pichia anomala. Process Biochemistry, April 2002, vol. 37, no. 9, p. 999-1004. [CrossRef] Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. |