
|
Assessment of genetic diversity and relationships among Osmanthus fragrans cultivars using AFLP markers Wang Jun Yuan1,2 · Yuan Ji Han1 · Mei Fang Dong1 · Fu De Shang*1 1 Key Laboratory of Plant Stress Biology,
College of Life Science, Henan University, Kaifeng 475004, PR China All of the authors contributed equally *Corresponding author: fudeshang@henu.edu.cn Financial support: This research was supported by the National Natural Science Funds in China (No. 30970176) and Innovation Scientists and Technicians Troop Construction Projects of Henan Province (No. 094100510018). Keywords: AFLP marker, cluster analysis, genetic diversity, Osmanthus fragrans.
This study was conducted to reveal genetic diversity among 100 Osmanthus fragrans cultivars using amplified fragment length polymorphism (AFLP) markers. Eight AFLP primer combinations produced a total of 443 polymorphic fragments with an average of 64 per primer combination. The percentage of polymorphic bands (86.81%), the resolving power (Rp) (32.71) and the PIC values (0.331) showed the efficiency of used primer combinations. The revealed AFLP makers were effective in distinguishing all the cultivars considered. Cluster analysis were performed to assess patterns of diversity among cultivars and showed the abundant genetic diversity. The overall distribution pattern of molecular variation suggested that 93.33% of the total genetic variance was within the identified groups and 6.67% of the genetic variation was among the identified groups. Our results showed that AFLP markers are useful for Osmanthus fragrans germplasm discrimination as well as for investigation of genetic diversity and variation. The information will facilitate germplasm identification, conservation and new cultivar development. Osmanthus fragrans, a species of Oleaceae family (Ômura et al. 2000), has been cultivated for more than 2,500 years in China. As a traditional horticultural plant, O. fragrans is valued for both its beauty and fragrance. The fragrance of O. fragrans contains as many as 35 potentially exploitable chemical substances (Wang et al. 2009). This plant also has medicinal value. According to Compendium of Materia Medica, the root of O. fragrans can be used to cure toothache. O. fragrans flowers are extensively used in Chinese cuisine. In 2004, the ISHS (International Society for Horticultural Science) appointed the Sweet Osmanthus branch of the Chinese Flower Association as International Cultivar Registration Authority (ICRA) for the genus Osmanthus. There are 122 registered O. fragrans cultivars (Xiang and Liu, 2008) growing mainly in the northern and middle subtropics between the Qinling and Nanling Mountains. The major centers of O. fragrans cultivation in China are Wu County of Jiangsu Province, Hangzhou of Zhejiang Province, Xianning of Hubei Province, Chengdu of Sichuan Province, and Guilin of Guangxi Province. In recent years, global warming has allowed a northward range expansion of O. fragrans. Since the mid-twentieth century, O. fragrans cultivars have been investigated and classified into Asiaticus and autumn divisions based on flowering season; the autumn division was further divided into three groups (Albus group, Luteus group, and Aurantiacus group) according to flower and leaf traits. Liu (1985) categorized O. fragrans cultivars into four groups (Asiaticus group, Albus group, Luteus group, and Aurantiacus group) on phenological and morphological traits. Liu’s classification procedure is the most detailed and has gained widespread acceptance. Using this procedure, 18 cultivars of the Asiaticus group, 57 of the Albus group, 36 of the Luteus group and 43 of the Aurantiacus group were identified in a nationwide investigation. Xiang and Liu (2008) published ‘An Illustrated Guide to Osmanthus fragrans Cultivars of China’, which recorded 122 O. fragrans cultivars and provided detailed characteristics of these cultivars. Genetic diversity study of O. fragrans on the molecular level has advanced significantly due to the rapid development of molecular biology. The results of a RAPD analysis of some Wuhan cultivars appeared to support the traditional morphology-based classification criteria, demonstrating high levels of genetic diversity of O. fragrans. Employing RAPD and ISSR-PCR techniques, several recent studies presented similar results. For example, Li et al. (2009) used the SRAP technique to analyze O. fragrans cultivars; the results indicate that many O. fragrans Albus cultivars are highly mutated, and some Luteus cultivars with deeper-colored flowers are closely related to certain members of the Aurantiacus group. Using Amplified fragment length polymorphism (AFLP) markers, Yan et al. (2009) analyzed some O. fragrans cultivars. Because of less cultivars used and only a few certified by International Cultivar Registration Authority (ICRA), the results of the previous studies are limited in reveal the genetic diversity of O. fragrans. We report here the genetic diversity of O. fragrans cultivars based on fluorescence-based AFLP makers. The two main objectives for germplasm management are: (1) to explore an effective mean of O. fragrans cultivars identification, (2) to establish of genetic database, which could provide molecular evidence for selection and breeding of the cultivars and genetic relationship in O. fragrans. Plant materials and DNA extraction This study used a total of 100 samples collected from the main cultivated land in China. The samples consisted of 13 cultivars in Asiaticus group, 31 cultivars in Albus group, 31 cultivars in Luteus group, and 25 cultivars in Aurantiacus group (Table 1). Total cellular DNA was extracted from frozen leaves sampled from each accession following the procedure described by Elameen et al. (2008). After purification, the DNA concentration was estimated using the spectrophotometer, then diluted to 100 ng/μl before used. AFLP analysis AFLP analysis was carried out based on the protocol described by Bonants et al. (2000). Genomic DNA (250 ng) was digested with PstI and MseI (2.5 U each) (Fermentas) in a final volume of 20 ml for 4 hrs at 37ºC and then the adapters were ligated to the digested DNA fragments. A preselective amplification using the primer pair PstI+G/MseI+C was performed using the following cycling parameters: 94ºC for 4 min; 30 cycles of 94ºC for 30 sec, 56ºC for 30 sec, 72ºC for 60 sec and 72ºC for 10 min. The 40-fold diluted preamplified DNA was amplified using the PstI+3 and MseI+3 primer pairs. The MseI+3 primers were labelled with a 6-FAM fluorescent dye. Eight primer combinations were used for the selective amplifications (Table 2). Amplification was conducted for 1 cycle of 95ºC for 30 sec, 65ºC for 40 sec and 72ºC for 60 sec, then lowering the annealing temperature by 0.7ºC decrements for 12 cycles; and then 23 cycles of 95ºC for 30 sec, 56ºC for 40 sec and 72ºC for 60 sec. The amplified products from selective amplifications were loaded and run on the ABI PRISM 377 DNA Sequencer (Applied Biosystems). Data were analyzed using GENESCAN 3.1 software. Data analysis The efficiency of each primer
combinations was estimated by the number of bands, and the polymorphic bands
were used to calculate the percentage of polymorphic bands (PPB). The estimation
of the resolving power (Rp) allowed the evaluation of the ability of the most
informative primers to differentiate between cultivars, Rp = ∑Ib; where:
Ib=1-(2 x|0.5-p|) and p is the
accessions proportion containing the I band (Ben Tamarzizt et al. 2009).
Moreover, the discriminating power of derived markers was calculated by the assessment
of the polymorphism information content (PIC) using the following formula: AFLP amplified fragments were scored as present (1) or absent (0) and entered into a binary qualitative data matrix. The presence/absence data matrix of the AFLP phenotypes was analyzed using the NTSYS-pc software package (Rohlf, 2000). A similarity matrix was constructed based on Dice’s coefficient. The similarity matrix was used to construct a dendrogram using the unweighted pair group method arithmetic average (UPGMA) to determine genetic relationships among the germplasm studied. The analysis of molecular variance (AMOVA) was performed. The variance components from the analysis were used to estimate the groups’ subdivisions (F-statistics) among cultivars from the four groups (FST). FST = VAP/ (VAP + VWP), where VAP is the variance among groups, VWP is the variance within groups (Peakall and Smouse, 2006). AFLP makers and polymorphism The 8 primer combinations, generating clear-cut AFLP profiles, resulted in a total of 514 different amplification products ranging from 50 to 500 bp in size with 443 polymorphic AFLP bands for 100 O. fragrans cultivars. The average of polymorphic bands per AFLP primer combination was 55.375 (Table 2) and each primer pair could distinguish all the cultivars considered. The largest number of polymorphic bands (66) was produced with primer combination PGAA/MCTG, and the least number of polymorphic bands (36) was detected using primer combination PGAC/MCAG. The percentage of polymorphic bands ranged from 80% for PGAC/MCAG to 95.65% for PGAA/MCTG primer combinations with a mean of 86.81% (Table 2). Thus, we assumed that all the tested primers were powerful to detect DNA polymophisms in O. fragrans cultivars. Estimates of the resolving power (Rp) showed a high rate of collective Rp, ranging from 16.18 to 39.26, with an average of 32.71 (Table 2). In addition, the polymorphism information content values varied from 0.245 to 0.393, 6 primer combinations used had a PIC exceeding 0.3, with a mean of 0.331, demonstrating a good discriminatory power of the primer combinations (Table 2). Therefore, data proved that AFLP was a powerful procedure to survey O. fragrans cultivars diversity. Cluster analysis among cultivars Based on the 443 AFLP markers, the level of similarity determined by the Dice’s coefficients ranged from 0.294 to 0.780 with a mean of 0.562, suggesting that the cultivars in this study were characterized by a great similarity at DNA level. The lowest Dice’s coefficient value of 0.294 has been scored between ‘Changyebizhu’ and ‘Jiaorong’. However, ‘Xiaoye Sijigui’ and ‘Tiannvsanhua’ were most similarly, with the highest Dice’s coefficient of 0.780. All the other ones have displayed different intermediate levels of genetic similarity. The dendrogram constructed using UPGMA cluster analysis (Figure 1) has clustered the 100 cultivars into six major clusters. The first cluster, labeled I, was dominated with 12 Asiaticus cultivars and 11 Albus cultivars. The second and the third clusters, II and III, consisted of 22 Aurantiacus and 14 Luteus cultivars. Cluster IV, V, VI were composed of the remaining cultivars, including 20 Albus cultivars, 17 Luteus cultivars, 3 Aurantiacus cultivars and 1 Asiaticus cultivar. Analysis of molecular variance (AMOVA) Within and between groups of cultivars, molecular variation was estimated using the AMOVA procedure. Total variance was partitioned into components due to differentiation within and among the four O. fragrans cultivar groups. The overall distribution pattern of molecular variation within the cultivars suggested that approximately 93.33% of the total variance could be accounted for by the within groups component of variance. The remaining 6.67% of the variation was founded among groups. The variance component was shown to be highly significant (P < 0.001). The number of permutations for significance testing was set at 1000 for analysis (Table 3). Information on genetic diversity and variation of germplasm is important for taxonomist, germplasm identification and new cultivar development (Demir et al. 2010). Morphological characters are often susceptible to environmental influences which may result in morphological differences between the same cultivar (Sarkhosh et al. 2009). Clear and detailed assessment of diversity within cultivars can be obtained by using molecular techniques. The variety discrimination through DNA profiles is useful for the protection and conservation of the genetic resources (Piña-Escutia et al. 2010). In the present work, AFLP technique has been used to analyze genetic diversity and relationships within the four groups. The majority of these 100 cultivars have never been considered for AFLP molecular marker studies. A total of 8 primer combinations were used in this study. Each primer combination was able to discriminate all the 100 cultivars studied. The high efficiency of primers in the genetic diversity was also supported by the measurement of resolving power and PIC value. The higher value of Rp and PIC belonged to the primer pairs PGAA/ MCTG and PGAC/ MCTC (39.26 and 0.393, respectively; Table 2), indicating a better distinction of the cultivars. These results also confirmed the utility of the Rp and PIC as a measure of the capacity of primer combination to discriminate among closely related individuals as pointed out by Prevost and Wilkinson, (1999) and Escandón et al. (2007). According to flower and leaf traits, O. fragrans cutilvars can be divided into four groups, Asiaticus group, Albus group, Leutus group and Aurantiacus group (Liu, 1985). The Albus group, caespitose shrub, flower mainly in spring and autumn, with paniculiform inflorescences in spring and cymose inflorescences in autumn. The other 3 groups are big shrub or tree, with cymose inflorescences and only flower in autumn. Flower color of the three groups differ substantially, with white, greenish white to yellowish white color (RHS-CC 1~8) in Albus group, yellow to golden yellow color (RHS-CC 9~20) in Leutus group and light orange-yellow to orange red (RHS-CC 21~30) in Aurantiacus group. In this study, the UPGMA cluster (Figure 1) showed the distribution of cultivars closeness among varieties with similar flower color. Cultivars in Albus group with light yellowish white flower color (18, 21, 22, 23, etc.) clustered together with Asiaticus group germplasms (Cluster I), showing a close genetic relationship between the two groups. Some Albus group cultivars (34, 35, 36, 38, etc.) and Luteus group cultivars (45, 46, 58, 59, etc.) clustered in cluster IV based on the similarity of the flower color (yellowish white to yellow). Cultivars in Leteus group with golden yellow flower color (61, 64, 67, 71, etc.) often clustered together with Aurantiacus group (Cluster III). Cluster V included 7 cultivars of three groups, showing the closeness among these cultivars. The other 2 clusters, II and VI were only consisted of Aurantiacus group cultivars and Albus group cultivars, respectively. The cluster analysis showed that there were close genetic relationships among cultivars with similar flower color and the flower color could be used as a good criterion of classification in discriminating O. fragrans germplasms. Our results are in agreement with some molecular marker study (Li et al. 2009; Yan et al. 2009), but only partially consistent with the traditional classification based on morphological characters. This might have been caused by that morphological characters are often under severe selection pressure which may result in morphological differences between the same cultivar, and AFLP markers are generally less susceptible to environmental impact. Dendrogram of four O. fragrans cultivar groups resulting from UPGMA cluster analysis (Figure 2), based on Dice’s coefficient, showed that Luteus group and Aurantiacus group had the closest genetic relationship, and Asiaticus group had a distant relationship with the other three groups. The results were consistent with those of a previous study (Li et al. 2009). The percentage of variations within group was, consequently, larger than that among groups. These findings suggested the presence of considerable cultivar variation among O. fragrans germplasm. The low divergence scored among the groups and the large variability detected within groups could be explained by the occurrence of gene flow in the natural populations from which cultivars originated and the reproduction mode. In fact, life form and breeding systems can have significant influences on the genetic variation and its partitioning (Salhi-Hannachi et al. 2005). Additionally, the FST values between cultivar groups ranged from 0.0280 to 0.1340 (Table 4). The greatest FST values were between Asiaticus and Luteus groups, and the lowest between Luteus and Aurantiacus groups. The FST value among overall groups was marginally lower (0.067), which indicated a lower degree of genetic differentiation. The current study demonstrated that AFLP is an effective approach to assess genetic diversity and was a powerful method to discriminate O. fragrans cultivars. The information may be useful for future germplasm identification, conservation and new cultivar development. BEN TAMARZIZT, H.; BARAKET, G.; BEN MUSTAPHA, S.; MARRAKCHI, M.; TRIFI, M. and SALHI-HANNACHI, A. (2009). Genetic relatedness among Tunisian plum cultivars by random amplified polymorphic DNA analysis and evaluation of phenotypic characters. Scientia Horticulturae, vol. 121, no. 4, p. 440-446. [CrossRef] BONANTS, P.J.M.; HAGENAAR-DE WEERDT, M.; MAN IN’T VELD, W.A. and BAAYEN, R.P. (2000). Molecular characterization of natural hybrids of Phytophthora nicotianae and P. cactorum. Phytopathology, vol. 90, no. 8, p. 867-874. [CrossRef] DEMIR, K.; BAKIR, M.; SARIKAMIŞ, G. and ACUNALP, S. (2010). Genetic diversity of eggplant (Solanum melongena) germplasm from Turkey assessed by SSR and RAPD markers. Genetics and Molecular Research, vol. 9, no. 3, p. 1568-1576. [CrossRef] ELAMEEN, A.; FJELLHEIM, S.; LARSEN, A.; ROGNLI, O.A.; SUNDHEIM, L.; MSOLLA, S.; MASUMBA, E.; MTUNDA, K. and KLEMSDAL, S. (2008). Analysis of genetic diversity in a sweet potato (Ipomoea batatas L.) germplasm collection from Tanzania as revealed by AFLP. Genetic Resources Crop Evolution, vol. 55, no. 3, p. 397-408. [CrossRef] ESCANDÓN, A.S.; ZELENER, N.; DE LA TORRE, M.P. and SOTO, S. (2007). Molecular identification of new varieties of Nierembergia linariaefolia (Graham), a native Argentinean ornamental plant. Journal of Applied Genetics, vol. 48, no. 2, p. 115-123. LI, M.; HOU, X.L. and HAO, R.M. (2009). Analysis of genetic relationships of Osmanthus fragrans based on SRAP markers. Acta Horticulturae Sinica, vol. 36, no. 11, p. 1667-1675. LIU, Y.L. (1985). Preliminary study of sweet osmanthus in Nanjing area. Journal of Nanjing Forestry University, vol. 1, p. 30-37. MUTHUSAMY, S.; KANAGARAJAN, S. and PONNUSAMY, S. (2008). Efficiency of RAPD and ISSR markers system in accessing genetic variation of rice bean (Vigna umbellata) landraces. Electronic Journal of Biotechnology, vol. 11, no. 3. [CrossRef] ÔMURA, H.; HONDA, K. and HAYASHI, N. (2000). Floral scent of Osmanthus fragrans discourages foraging behavior of cabbage butterfly, Pieris rapae. Journal of Chemical Ecology, vol. 26, no. 3, p. 655-666. [CrossRef] PEAKALL, R. and SMOUSE, P.E. (2006). GENALEX6: Genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes, vol. 6, no. 1, p. 288-295. [CrossRef] PIÑA-ESCUTIA, J.L.; VAZQUEZ-GARCIA, L.M. and ARZATE-FERNANDEZ, A.M. (2010). Variety discrimination of Tigridia pavonia (L.f.) DC. assessed by different length RAPD primers. Electronic Journal of Biotechnology, vol. 13, no. 4. [CrossRef] PREVOST, A. and WILKINSON, M.J. (1999). A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. TAG Theoretical and Applied Genetics, vol. 98, no. 1, p. 107-112. [CrossRef] ROHLF, F.J. (2000). NTSYS-pc, Numerical Taxonomy and Multivariate Analysis System, Version 2.1 Exeter Pulications, New York, USA. SALHI-HANNACHI, A.; CHATTI, K.; MARS, M.; MARRAKCHI, M. and TRIFI, M. (2005). Comparative analysis of genetic diversity in two Tunisian collections of fig cultivars based on random amplified polymorphic DNA and Inter simple sequence repeats fingerprints. Genetic Resources and Crop Evolution, vol. 52, no. 5, p. 563-573. [CrossRef] SARKHOSH, A.; ZAMANI, Z.; FATAHI, R. and RANJBAR, H. (2009). Evaluation of genetic diversity among Iranian soft-seed pomegranate accessions by fruit characteristics and RAPD markers. Scientia Horticulturae, vol. 121, no. 3, p. 313-319. [CrossRef] UZUN, A.; YESILOGLU, T.; AKA-KACAR, Y.; TUZCU, O. and GULSEN, O. (2009). Genetic diversity and relationships within Citrus and related genera based on sequence related amplified polymorphism markers (SRAPs). Scientia Horticulturae, vol. 121, no. 3, p. 306-312. [CrossRef] WANG, L.M.; LI, M.T.; JIN, W.W.; LI, S.; ZHANG, S.Q. and YU, L.J. (2009). Variations in the components of Osmanthus fragrans Lour. essential oil at different stages of flowering. Food Chemistry, vol. 114, no. 1, p. 233-236. [CrossRef] XIANG, Q.B. and LIU, Y.L. (2008). An illustrated monograph of the sweet osmanthus cultivars in China. Zhejiang Science and Technology Publishing House, 377 p. ISBN978-7-5341-3185-1. YAN, X.Y.; XIAO, B.L.; HAN, Y.J.; YUAN, W.J. and SHANG, F.D. (2009). AFLP analysis of genetic relationships and diversity of some Chinese Osmanthus fragrans cultivars. Life Science Journal, vol. 6, no. 2, p. 11-16. Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. |
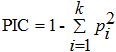 , where Pi is the frequency
of the ith allele in the set of genotypes investigated (
, where Pi is the frequency
of the ith allele in the set of genotypes investigated (