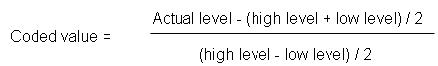|
Optimization of β-galactosidase production using Kluyveromyces lactis NRRL Y-8279 by response surface methodology Seval Dagbagli Yekta Goksungur* *Corresponding author Financial support: This research was supported by the State Planning Organization (Turkey) through Project no: DPT.2005K120570 and Scientific and Technical Research Council of Turkey (TÜBİTAK) through Project no: TOVAG 104 O 270. Keywords: β-galactosidase, Kluyveromyces lactis, response surface methodology, shake flask culture, synthetic medium.
This paper investigates the production and optimization of β-galactosidase enzyme using synthetic medium by Kluyveromyces lactis NRRL Y-8279 in shake flask cultures. Among the different cell disintegration methods used, the highest specific activity was obtained when the cells were permeabilized using isoamyl alcohol. Response surface methodology was used to investigate the effects of four fermentation parameters (agitation speed, pH, initial substrate concentration and incubation time) on β-galactosidase enzyme production. Results of the statistical analysis showed that the fit of the model was good in all cases. Maximum specific enzyme activity of 4218.4 U g-1 was obtained at the optimum levels of process variables (pH 7.35, agitation speed 179.2 rpm, initial sugar concentration 24.9 g l-1 and incubation time 50.9 hrs). The response surface methodology was found to be useful in optimizing and determining the interactions among process variables in β-galactosidase enzyme production.
The enzyme β-galactosidase (lactase, EC 3.2.1.23) catalyzes the hydrolysis of lactose to glucose and galactose. This enzyme is industrially important because it can be used to avoid lactose crystallization in sweetened, condensed and frozen dairy products such as ice cream and condensed milk and solve problems associated with whey utilization and disposal. In addition, β-galactosidase is used to avoid the problems of lactose intolerance by individuals who are deficient in lactase (Artolozaga et al. 1998). New applications for β-galactosidase, such as in the production of biologically active galacto-oligosaccharides, have also been reported in the literature (Boon et al. 2000; Albayrak and Yang, 2002). Commercial β-galactosidases are produced from yeasts such as Kluyveromyces lactis and Kluyveromyces marxianus (formerly known as Kluyveromyces fragilis and Saccharomyces fragilis), and moulds such as Aspergillus niger and Aspergillus oryzae (Shaikh et al. 1997; Santos et al. 1998). β-galactosidases produced by yeasts are the most employed for the treatment of milk, sweet whey and neutral pH dairy products since their optimum pH is between 6.5-7.0 (Santos et al. 1998). The activity and stability of enzymes is influenced by the type of strain, cultivation conditions (temperature, pH, aeration, agitation, incubation time) and the growth medium composition (particularly carbon and nitrogen sources) (Schneider et al. 2001; Jurado et al. 2004; Tari et al. 2007). Several papers have been published (Chen et al. 1992; Fiedurek and Szczodrak, 1994; Bojorge et al. 1999; Furlan et al. 2000; Furlan et al. 2001) reporting the optimization of a variety of culture conditions for the production of β-galactosidase by Kluyveromyces marxianus. The optimization studies done by varying one parameter while keeping the others at constant level do not reflect the interaction effects among these variables employed and this kind of optimization studies do not depict the net effect of the various factors on the enzyme activity. In order to overcome this major problem, optimization studies are done using response surface methodology (RSM) which is a mathematical and statistical technique widely used to determine the effects of several variables and to optimize different biotechnological processes (He and Tan, 2006). RSM has been extensively applied to optimize culture medium and other process parameters for the production of lipase (He and Tan, 2006; Liu et al. 2006), tannase (Battestin and Macedo, 2007), α-amylase (Uma Maheswar Rao and Satyanarayana, 2007), β-cyclodextrin glucanotransferase (Ibrahim et al. 2005), dextran dextrinase (Naessens et al. 2004) and chitinase (Nawani and Kapadnis, 2005). Chen et al. (1992) studied the optimization of β-galactosidase production using a statistical and mathematical approach. These authors used a central composite design to optimize the medium composition for lactase production by Kluyveromyces marxianus and found the optimal medium composition as 80.6 g l-1 lactose, 107.7 g l-1 corn steep liquor, 4.1 g l-1 glucose and 9.6 g l-1 glycerol. Becerra and González Siso (1996) used a full-factorial design to optimize β-galactosidase production by Kluyveromyces lactis in solid-state fermentations on corn grits or wheat bran moistened with deproteinized milk whey. No previous work has used RSM or other statistical techniques to optimize cultivation conditions and to determine the interactions among pH, agitation speed, incubation time and initial sugar concentration in β-galactosidase production. The present study examined β-galactosidase production from synthetic medium containing lactose by Kluyveromyces lactis NRRL Y-8279. The effects of cell disruption methods on β-galactosidase activity were determined. RSM was used to optimize fermentation parameters to obtain maximum enzyme activity. Four factors (pH, agitation speed, initial sugar concentration and incubation time) considered to have significant impact on enzyme production were selected as parameters for optimization studies. This study is the first detailed work on the use of response surface methodology for the optimization of fermentation parameters in β-galactosidase production using K. lactis in shake flask cultures. Kluyveromyces lactis NRRL Y-8279 used throughout this study was kindly supplied by the U.S. Department of Agriculture, National Center for Agricultural Utilization Research. The strain is maintained on Yeast-Malt agar slants stored at 4ºC and transferred monthly to fresh slants incubated at 30ºC for 2 days. Two wire loops of culture were taken from fresh slants and transferred to 250 ml Erlenmayer flasks containing 50 ml of culture broth which had been sterilized at 121ºC for 15 min. Medium composition of agar slants was as follows (in g l-1): glucose, 10; yeast extract, 3; malt extract, 3; peptone, 5; agar, 20 (pH = 6.0 ± 0.2). Medium composition for inoculum and fermentation were (in g l-1): lactose, 30; yeast extract, 1; K2HPO4, 2; NH4H2PO4, 1; (NH4)2HPO4, 1; MgSO4 x 7 H2O, 0.1 and pH was adjusted to 7.0 (unless otherwise stated) (Fiedurek and Szczodrak, 1994). All chemicals used in this study were of analytical grade. Fermentations were carried out batchwise in 250 ml flasks using 50 ml of fermentation medium sterilized at 121ºC for 15 min. The flasks were inoculated with the pre-inoculum volume (2%) to give an initial cell concentration between 0.05-0.1 g l-1. The cultures were incubated at 30ºC for 48 hrs in a rotary shaker incubator (Gerhardt Laboshake LS 2/5 RO 2/5, Bonn, Germany) operated at 200 rpm. The culture medium and growth conditions used for both inoculation and fermentation were the same as described above. The levels of pH, agitation speed, initial sugar concentration and incubation time used in the optimization studies by RSM are given in Table 1. β-galactosidase enzyme from K. lactis is an intracellular enzyme. Different chemical and mechanical methods were used to permeabilize and/or disrupt K. lactis cells prior to the assay of intracellular β-galactosidase. Hence, the effect of these methods on enzyme activity was determined. Isoamyl alcohol. A volume of resuspended cells in 0.2 M phosphate buffer (pH = 6.5) amounting to 10-20 mg of dry cell mass was mixed with 5 ml isoamyl alcohol and diluted up to 25 ml with 0.2 M phosphate buffer, pH 6.5. The mixture was shaken for 15 min at room temperature to make the cell envelopes permeable and used for the enzyme assay (Barberis and Gentina, 1998). Vortexing cells with glass beads. Cell suspensions were vortexed at 4ºC with glass beads (1 mm diameter) in a 30 ml glass tube. Vortexing continued for 10 min and the resulting mixture was centrifuged at 4000 x g. The supernatant was used for the enzyme assay (Song and Jacques, 1997). Liquid nitrogen. Biomass was placed in mortar and ground with liquid nitrogen and centrifuged at 4000 x g for 20 min. The supernatant was used for the enzyme assay (Liu et al. 2001). Triton X-100. Cell suspensions were shaken in presence of 0.04% Triton X-100 in a shaker at 200 rpm for 1 hr at 30ºC and centrifuged at 4000 x g for 20 min. The supernatant was used for the enzyme assay (Fiedurek and Szczodrak, 1994). Sodium dodecyl sulfate (SDS). Cell suspensions were shaken in presence of 0.04% SDS in a shaker at 200 rpm for 1 hr (30ºC) and centrifuged at 4000 x g for 20 min. The supernatant was used for the enzyme assay (Fiedurek and Szczodrak, 1994). Sonication. The cell suspensions were sonicated on ice in glass tubes using a Branson Sonic Power Sonicator (48 Bransonic Power, 40 W, 30 sec with 30 sec cooling periods) for 4 min. Then, they were centrifuged at 4000 x g for 20 min and the supernatant was used for measuring β-galactosidase activity (Song and Jacques, 1997). The method described by Food Chemical Codex (1993) was used with some modifications for β-galactosidase activity. The substrate for the enzyme was 2.5 mg ml-1 of ο-nitrophenol-β-D-galactopyranoside (ONPG, Sigma, N-1127), which was prepared in 0.2 M phosphate buffer, pH = 6.5. 1 ml of the crude permeabilized cell suspension with isoamyl alcohol (unless otherwise stated) was incubated with 4 ml of the above substrate mixture (preheated to 37ºC) at 37ºC for 15 min. The reaction was terminated by adding 1 mL of 10% sodium carbonate. The liberated o-nitrophenol (ONP) was measured spectrophotometrically at 420 nm. ONP concentration was calculated using an extinction coefficient of 4.2371 ml μmol-1 cm-1. One unit of enzyme activity was defined as the enzyme quantity that liberated 1 μmol of o-nitrophenol per minute under the assay conditions. Specific activity (U g-1) was defined as the number of lactase units per gram of cell biomass. Specific β-galactosidase activity was used throughout the study to obtain a common basis for comparisons. Volumetric enzyme activity values were only given in determining the kinetics of enzyme production. The biomass concentration was measured gravimetrically as dry cell mass (g l-1) by centrifuging 20 ml of original culture broth at 4000 x g for 20 min followed by drying the cells at 80ºC overnight. Lactose was measured using the 3,5-dinitrosalicylic acid (DNS) method (Miller, 1959). All experiments were carried out in triplicate and the values reported are the mean of three such experiments in which 3-5% variability was observed. Experimental design and statistical analysis The statistical analysis of the data was performed using Minitab Statistical Software (Release 13.20). Details of response surface methodology can be found elsewhere (Myers and Montgomery, 1995). The levels of factors used in the experimental design are listed in Table 1. The data of the factors were chosen after a series of preliminary experiments. Thirty one experiments were conducted using a central composite routable design for the study of four factors each at five levels (Table 2). The levels were –a, -1, 0, +1, +a where a = 2n/4, n was the number of variables and 0 corresponded to the central point. The actual level of each factor was calculated using the following equation (Myers and Montgomery, 1995).
Response surface model was fitted to the response variable, namely specific β-galactosidase activity (U g-1). The second order response function for four quantitative factors is given by Equation [1]: Y = β0 + β1X1 + β2X2 + β3X3 + β4X4 + β11X12 + β22X22 + β33X32 + β44X42 + β12X1X2 + β13X1X3 + β14X1X4 + β23X2X3 + β24X2X4 + β34X3X4 [1] where X1, X2, X3 and X4 represent the levels of the factors according to Table 1 and β0, β1,......, β34 represent coefficient estimates with β0 having the role of a scaling constant. Effect of cell disintegration methods on specific enzyme activity Cell disruption methods can be roughly divided into those which produce cell leakage and disruption in response to the application of shear forces (mechanical methods) and those which produce digestion or permeabilization of the cell envelopes by the action of chemicals (chemical methods) (Numanoglu and Sungur, 2004). In permeabilization, the cell envelope is altered to allow small molecules, such as substrates, products or coenzymes to cross freely. The permeabilizing agent disrupts the membrane structures to allow the passive passage of low molecular weight solutes in and out of cells. Organic solvents as permeabilizing agents act on the cell membranes by decreasing the phospholipids content (Panesar et al. 2007). In this study, among all the mechanical and chemical cell disintegration methods tested, the highest specific enzyme activity was obtained when K. lactis cells were treated with isoamyl alcohol prior to the assay of β-galactosidase. After permeabilization of cell walls of K. lactis with isoamyl alcohol, ONPG diffused into the cells and was hydrolyzed by the β-galactosidase enzyme entrapped within the cell. Cell disintegration is an important procedure applied in the production of intracellular β-galactosidase enzyme from K. lactis. Different mechanical and chemical disruption methods were applied to the cell mass at the end of fermentation. The results indicated that the specific activity of β-galactosidase enzyme is dependent on the cell disintegration method used. As seen in Figure 1, highest specific enzyme activity (3416.6 U g-1) was obtained when the cells were permeabilized using isoamyl alcohol. Similar specific enzyme activity was obtained by disrupting the cells mechanically using glass beads (3038.9 U g-1). Lower specific enzyme activities were found when Triton X-100 (1888.8 U g-1), liquid nitrogen (1199.4 U g-1) and SDS (964.3 U g-1) were used. The lowest enzyme activity (152.6 U g-1) was obtained with the sonication method. Several studies have been reported on the permeabilization of microbial cells by organic solvents (Flores et al. 1994; Kondo et al. 2000; Numanoglu and Sungur, 2004; Panesar et al. 2007; Park et al. 2007). Flores et al. (1994) studied the permeabilization of K. lactis cells by chloroform, toluene and ethanol in relation to β-galactosidase activity. They found that the performance of those solvents was dependent on the incubation time, the incubation temperature and the concentration of both cells and solvents. Panesar et al. (2007) used ethanol to permeabilize K. marxianus cells to overcome the problem of enzyme extraction and poor permeability of cell membrane in the production of lactose-hydrolyzed milk. Park et al. (2007) investigated the use of five different solvents (dimethyl sulfoxide, petroleum ether, acetone, chloroform and hexane) to disrupt the cell wall of freeze-dried Rhodotorula glutinis cells to extract intracellular carotenoid pigments. Kondo et al. (2000) permeabilized flocculent yeast Saccharomyces cerevisiae with isopropyl alcohol and ethanol under various conditions. Other chemicals used to increase the membrane permeability of yeast cells were toluene-ethanol mixture, cethyltrimethyl-ammonium bromide, ethyl ether, digitonin, Triton X-100 and hexamethylenediamine (Kondo et al. 2000). Mechanical methods such as sonication, high-pressure homogenizer or bead mills have been traditionally used for the disruption of microbial cells (Geciova et al. 2002). However such techniques suffer from drawbacks, including the production of fine cell debris, high capital investment cost and long processing time (Park et al. 2007). The method used should be robust enough to disrupt cell envelopes efficiently but gentle enough to preserve enzyme activity (Numanoglu and Sungur, 2004). Several papers have been published on the use of mechanical methods for the release of intracellular β-galactosidase enzyme. Numanoglu and Sungur (2004) compared chemical (toluene, ethanol-chloroform) and physical (glass bead mill) methods to permeabilize β-galactosidase producing K. lactis cells and found that the physical method was better because of the probable risk of chemical toxicity accompanied with the chemical methods. Fiedurek and Szczodrak (1994) used three types of extraction methods to release the β-galactosidase enzyme from K. fragilis cells and found that the highest yield was obtained by mechanical disintegration of cells by mortar and pestle. In this study, among the mechanical methods tested for cell disruption, the highest specific enzyme activity was obtained by mechanical disruption of K. lactis cells with glass beads. In our study, sonication was found to be unsuitable for cell disruption. This might be because of the incomplete disruption of the K. lactis cell wall since lactase is a large enzyme and complete disruption of the cell and its cell wall is necessary to liberate the enzyme from the cell debris. Another reason for obtaining low enzyme activity units in sonication might be the significant heat emerging from the absorption of sonication energy into suspensions, probably leading to enzyme inactivation. Therefore, good temperature control is necessary throughout the disruption process. Parallel to the findings of this study, Bury et al. (2001) studied the disruption of Lactobacillus delbrueckii ssp. bulgaricus 11842 cells to compare sonication, high-pressure homogenization and bead milling for the release of β-galactosidase enzyme. They found that bead milling and high-pressure homogenizer were comparable methods while laboratory scale sonication was the least effective method on the release of β-galactosidase enzyme. Contrarily, Berger et al. (1995) studied two physical disruption methods to optimize the recovery of intracellular enzymes from Thermus species and found that the sonication method was superior to the glass-bead method in the recovery of the total thermophilic β-galactosidase activity. Agrawal and Pandit (2003) studied cell disruption of S. cerevisiae with ultrasonication for the release of intracellular α-glucosidase enzyme and optimized the pH of the disruption medium for both unit activity and specific activity of the enzyme. Iida et al. (2008) investigated the release of intracellular protein from yeast cells by the ultrasonic action and studied the effect of factors such as the concentration of yeast cells, temperature, ultrasound power, types of sonicator and the superposition with the mechanical mixing. In our sonication system, optimizing the operational parameters such as time and power might have helped to achieve less enzyme inactivation and higher enzyme activity. The completeness of cell disruption by sonication could have been determined by using a microscope and/or by measuring the release of nucleic acids from the disrupted cells. Kinetics of β-galactosidase production from K. lactis NRRL Y-8279 The kinetics of K. lactis growth and production of β-galactosidase enzyme in the shake flask batch culture were examined by growing the cells in the synthetic medium containing 30 g l-1 of initial lactose. As seen in Figure 2, specific enzyme activity curve exhibited two distinct sharp points at the beginning and at the end of the growth phase. The specific activity remained low at the beginning of the cultivation and then increased giving a peak at the early exponential phase. During the exponential growth phase, specific activity increased reaching a maximum at the early stationary phase and decreased continuously throughout the stationary growth phase. The maximum specific activity (3416.6 U g-1) was attained at the end of 48 hrs. The reason for obtaining two distinct points on specific activity curve might be attributed to the hydrolysis of lactose to glucose by the β-galactosidase enzyme synthesized in the initial stages of fermentation. The glucose produced might have repressed the enzyme synthesis. After exhaustion of glucose, the microorganism might have synthesized more β-galactosidase in an effort to utilize lactose in the medium leading to a second peak of enzyme activity. It was observed from the volumetric enzyme activity curve (Figure 2) that the enzyme production was associated with the biomass production and the maximum volumetric activity (14106 U l-1) was obtained at the 48th hrs of fermentation. However, volumetric activity decreased drastically at the stationary growth phase of the organism most probably due to the unstability of the intracellular enzyme, presence of proteases and changes in the cell wall during the stationary phase as well as the enzymatic inhibition of the hydrolysis products. The decrease of β-galactosidase activity after 48 hrs indicates that the operation should be stopped after the stationary phase has been reached. Hence, the fermentation time seems to be an important parameter that should be considered in optimization studies using RSM to achieve maximum enzyme activity. Maximum specific (189.83 U g-1 h) and volumetric (293.88 U l-1 h-1) productivity values were obtained at the end of 8 and 48 hrs, respectively. As expected, the concentration of residual sugars decreased during the fermentation and almost complete sugar depletion was observed in the culture medium. The residual sugar concentration in culture medium was 1.1 g l-1. In all culture systems, the pH decreased slightly during fermentation and decreased from an initial value of 7.0 to 5.0-5.3 at the end of 48 hrs. Similar activity profile curves were reported in the literature (Rech et al. 1999; Martins et al. 2002; Rajoka et al. 2003). Rech et al. (1999) studied β-galactosidase production from sweet cheese whey by using two different strains of K. marxianus and found that the specific activity curves exhibited two distinct sharp points at the beginning and at the end of the growth phase. They also stated that volumetric β-galactosidase activity followed the pattern of the growth curve. Martins et al. (2002) studied the production of β-galactosidase enzyme from K. marxianus and observed that the specific enzyme activity increased and attained a maximum after 4 hrs of incubation. They found a second phase of high specific activity after 16 hrs of incubation where the culture entered the stationary phase of growth. Rajoka et al.(2003) studied the production of β-galactosidase enzyme from K. marxianus in shake flask cultures and reported that the bulk β-galactosidase production reached a maximum activity after 30-40 hrs and the enzyme production was apparently growth associated. Optimization of β-galactosidase enzyme production Many authors stated the importance of factors such as pH, aeration, substrate concentration and fermentation time in β-galactosidase enzyme production (Chen et al. 1992; Fiedurek and Szczodrak, 1994; Shaikh et al. 1997; Rech et al. 1999; Furlan et al. 2000; Furlan et al. 2001; Fekete et al. 2002; Martins et al. 2002; Rajoka et al. 2003; Hsu et al. 2005; Konsoula and Liakopoulou-Kyriakides, 2007). Our preliminary experiments also showed that the agitation speed, pH, initial substrate concentration and incubation time influenced the production of β-galactosidase enzyme by K. lactis NRRL Y-8279. Thus, a central composite design was used to determine the optimum levels of these parameters leading to a maximum β-galactosidase enzyme synthesis. The levels of these factors (Table 1) used in the optimization studies by RSM were determined by preliminary experiments. The effect of the four previously mentioned variables, each at five levels, and their interactions on β-galactosidase enzyme synthesis have been determined by carrying out thirty one experiments given by the model (Table 2). Analysis of variance (ANOVA) for specific enzyme activity is presented in Table 3. The analysis gives the value of the model and determines the requirement of a more complex model with a better fit. As shown in Table 3, R2 was 0.998 indicating that the model as fitted explained 99.8% of the variability in specific enzyme activity. F-test for regression was significant at a level of 5% (P < 0.05) indicating that the model is fit and can adequately explain the variation observed in enzyme synthesis with the designed levels of the factors. If the F-test for lack of fit is significant, then a more complicated model is needed to fit the data. As seen in Table 3, the lack of fit (0.163) was not significant at the 5% level (P > 0.05) indicating that the experimental data obtained fitted well with the model. These results show that the model chosen can satisfactorily explain the effects of pH, agitation speed, initial substrate concentration and incubation time on β-galactosidase production by K. lactis NRRL Y-8279in shake flask cultures. Thirty one experiments were carried out from the design and by applying multiple regression analysis on the experimental data; the following second order polynomial equation was found to explain β-galactosidase production by K. lactis. Y = - 20297.9 + 4944.3 X1 + 28.319 X2 + 180.239 X3 + 57.784 X4 - 359.041 X12 - 0.06912 X22 - 2.66287 X32 - 0.84937 X42 - 4.46805 X1X3 + 8.68165 X1X4+ 0.087861 X2X3 - 0.11267 X2X4 - 0.60017 X3X4 [2] where X1, X2, X3 and X4 are the actual levels of factors shown in Table 1. Regression analysis (Table 4) of the experimental data showed that pH, agitation speed, initial substrate concentration and incubation time had positive linear effects on enzyme synthesis (P< 0.05). Probability (P) values were used as a tool to check the significance of each of the coefficients. The smaller the magnitude of P value, the more significant was the correlation with the corresponding coefficient. Among the four factors tested, pH had the highest impact on specific enzyme activity as given by the highest linear coefficient (4944.3), followed by initial substrate concentration (180.239), incubation time (57.7840) and agitation speed (28.3190). These factors also showed significant negative quadratic effects on enzyme production indicating that specific enzyme activity increased as the level of these factors increased and decreased as the level of these parameters increased above certain values. Interaction between these parameters was also significant. The interactions between pH-initial substrate concentration, pH-incubation time, agitation- initial substrate concentration, agitation-incubation time, initial substrate concentration-incubation time were significant as shown by low P values (P < 0.05) for interactive terms. But the interaction between pH-agitation speed was found to be insignificant as given by P value above 0.05. Hence this term was excluded from the polynomial Equation [2] used for this model. Figure 3 shows the contour plots of specific enzyme activity for each pair of factors by keeping the other two factors constant at its middle level. Maximum specific enzyme activity was obtained at middle level of each pair of factors at a constant middle level of the other two factors. Further increase in these factors above the middle level (pH: 7.3, agitation speed: 180 rpm, initial substrate concentration: 25 g l-1, incubation time: 50 hrs) showed decrease in specific enzyme activity. In order to determine the maximum specific enzyme activity corresponding to the optimum levels of pH, agitation speed, initial substrate concentration and incubation time, a second order polynomial model was used to calculate the values of these variables. The fitting of the experimental data to Equation [2] allowed the determination of the levels of pH (X1 = 7.35), agitation speed (X2 = 179.2 rpm), initial sugar concentration (X3 = 24.9 g l-1) and incubation time (X4 = 50.9 hrs) giving a maximum specific enzyme activity of 4113.1 U g-1. The above data optimizes β-galactosidase enzyme production from synthetic medium containing lactose by K. lactis NRRL Y-8279 in shake flask culture. The final fermentation was performed using synthetic medium containing 5 g l-1 yeast extract with the optimized levels of pH (7.35), agitation speed (179.2 rpm), initial sugar concentration (24.9 g l-1) and incubation time (50.9 hrs) given by the model. Maximum specific enzyme activity (4218.4 U g-1) which was slightly higher than the value given by the model was obtained on the 50.9th hour of fermentation. The maximum enzyme productivity obtained was 339.8 U l-1 h-1. In this study, β-galactosidase enzyme was produced by K. lactis NRRL Y-8279 using a synthetic medium containing lactose. Different chemical and mechanical methods were used for the disintegration of K. lactis cells and the highest specific enzyme activity (3416.6 U g-1) was obtained when the cells were permeabilized using isoamyl alcohol. The maximum specific enzyme activity of 3737.2 U g-1 was obtained at the end of 48 hrs. RSM was used to determine the effects of four important factors (agitation speed, pH, initial substrate concentration and incubation time) on β-galactosidase enzyme production. Linear, quadratic and interaction effects of these variables on specific enzyme activity were determined. The model generated in this study by RSM satisfied all the necessary arguments for its use in the optimization. By fitting the experimental data to a second order polynomial equation, the optimum levels of pH (7.35), agitation speed (179.2 rpm), initial sugar concentration (24.9 g l-1) and incubation time (50.9 hrs) were determined. Using the optimum levels of fermentation parameters, a maximum specific enzyme activity of 4218.4 U g-1 was obtained. This article provides a detailed study that used statistical analysis to determine the optimum levels and interactions among the above mentioned parameters in β-galactosidase enzyme production from K. lactis. The parameters studied in this article also influence the enzyme synthesis in a bioreactor system and further studies will be focused on the scale-up of enzyme production and the optimization of fermentation parameters in a stirred tank bioreactor. AGRAWAL, P.B. and PANDIT, A.B. Isolation of α-glucosidasefrom Saccharomyces cerevisiae: cell disruption and adsorption. Biochemical Engineering Journal, July 2003, vol. 15, no. 1, p. 37-45. [CrossRef] ALBAYRAK, N. and YANG, S.T. Production of galacto-oligosaccharides from lactose by Aspergillus oryzae β-galactosidase immobilized on cotton cloth. Biotechnology and Bioengineering, January 2002, vol. 77, no. 1, p. 8-19. ARTOLOZAGA, M.J.; JONAS, R.; SCHNEIDER, A.L.; FURLAN, S.A. and CARVALHO-JONAS, M.F. One step partial purification of β-D-galactosidase from Kluyveromyces marxianus CDB 002 using STREAMLINE-DEAE. Bioseparation, May 1998, vol. 7, no. 3, p. 137-143. [CrossRef] BARBERIS, S. and GENTINA, J.C. Effect of dissolved oxygen level on lactase production by Kluyveromyces fragillis. Journal of Chemical Technology and Biotechnology, September 1998, vol. 73, no. 1, p. 71-73. [CrossRef] BATTESTIN, V. and MACEDO, G.A. Tannase production by Paecilomyces variotii. Bioresource Technology, July 2007, vol. 98, no. 9, p. 1832-1837. [CrossRef] BECERRA, M. and GONZÁLEZ SISO, M.I. Yeats β-galactosidase in solid-state fermentation. Enzyme and Microbial Technology, July 1996, vol. 19, no. 1, p. 39-44. [CrossRef] BERGER, J.L.; LEE, B.H. and LACROIX, C. Identification of new enzyme activities of several strains of Thermus species. Applied Microbiology and Biotechnology, December 1995, vol. 44, no. 1-2, p. 81-87. [CrossRef] BOJORGE, N.; VALDMAN, B.; ACEVEDO, F. and GENTINA, J.C. A semi-structured model for the growth and β-galactosidase production by fed-batch fermentation of Kluyveromyces marxianus. Bioprocess and Biosystem Engineering, 1999, vol. 21, no. 4, p. 313-318. [CrossRef] BOON, M.A.; RIET VAN’T, K. and JANSSEN, A.E.M. Enzymatic synthesis of oligosaccharides: product removal during a kinetically controlled reaction. Biotechnology and Bioengineering, November 2000, vol. 70, no. 4, p. 411-420. BURY, D.; JELEN, P. and KALÁB, M. Disruption of Lactobacillus delbrueckii ssp. bulgaricus 11842 cells for lactose hydrolysis in dairy products: a comparison of sonication, high-pressure homogenization and bead milling. Innovative Food Science & Emerging Technologies, March 2001, vol. 2, no. 1, p. 23-29. [CrossRef] CHEN, K.C.; LEE, T.C. and HOUNG, J.Y. Search method for the optimal medium for the production of lactase by Kluyveromyces fragillis. Enzyme and Microbial Technology, August 1992, vol. 14, no. 8, p. 659-664. [CrossRef] FEKETE, E.; KARAFFA, L.; SÁNDOR, E.; SEIBOTH, B.; BIRÓ, S.; SZENTIRMAI, A. and KUBICEK, C.P. Regulation of formation of the intercellular β-galactosidase activity of Aspergillus nidulans. Archives Microbiology, December 2002, vol. 179, no. 1, p. 7-14. [CrossRef] FIEDUREK, J. and SZCZODRAK, J. Selection of strain, culture conditions and extraction procedures for optimum production of β-galactosidase from Kluyveromyces fragillis. Acta Microbiologica Polonica, 1994, vol. 43, no. 1, p. 57-65. FLORES, M.V.; VOGET, C.E. and ERTOLA, R.J.J. Permeabilization of yeast cells (Kluyveromyces lactis) with organic solvents. Enzyme and Microbial Technology, April 1994, vol. 16, no. 4, p. 340-346. [CrossRef] FOOD CHEMICALS CODEX. Lactase (β-galactosidase) activity, 3rd ed. National Academy Press, Washington, DC, 1993, 491 p. FURLAN, S.A.; SCHNEIDER, A.L.S.; MERKLE, R.; CARVALHO-JONAS, M.F. and JONAS, R. Formulation of a lactose-free, low-cost culture medium for the production of β-D -galactosidase by Kluyveromyces marxianus. Biotechnology Letters, April 2000, vol. 22, no. 7, p. 589-593. [CrossRef] FURLAN, S.A.; SCHNEIDER, A.L.S.; MERKLE, R.; CARVALHO-JONAS, M.F. and JONAS, R. Optimization of pH, temperature and inoculum ratio for the production of β-D -galactosidase by Kluyveromyces marxianus using a lactose free medium. Acta Biotechnologica, February 2001, vol. 21, no. 1, p. 57-64. [CrossRef] GECIOVA, J.; BURY, D. and JELEN, P. Methods for disruption of microbial cells for potential use in the dairy industry- a review. International Dairy Journal, 2002, vol. 12, no. 6, p. 541-553. [CrossRef] HE, Y.Q. and TAN, T.W. Use of response surface methodology to optimize culture medium for production of lipase with Candida sp. 99-125. Journal of Molecular Catalysis B: Enzymatic, December 2006, vol. 43, no. 1-4, p. 9-14. [CrossRef] HSU, C.A.; YU, R.C. and CHOU, C.C. Production of β-galactosidase by Bifidobacteria as influenced by various culture conditions. International Journal of Food Microbiology, October 2005, vol. 104, no. 2, p. 197-206. [CrossRef] IBRAHIM, H.M.; YUSOFF, W.M.W.; HAMID, A.A.; ILLIAS, R.M.; HASSAN, O. and OMAR, O. Optimization of medium for the production of β-cyclodextrin glucanotransferase using central composite desing (CCD). Process Biochemistry, February 2005, vol. 40, no. 2, p. 753-758. [CrossRef] IIDA, Y.; TUZIUTI, T.; YASUI, K.; KOZUKA, T. and TOWATA, A. Protein release from yeast cells as an evaluation method of physical effects in ultrasonic field. Ultrasonics Sonochemistry, September 2008, vol. 15, no. 6, p. 995-1000. [CrossRef] JURADO, E.; CAMACHO, F.; LUZÓN, G. and VICARIA, J.M. Kinetic models of activity for β-galactosidases: influence of pH, ionic concentration and temperature. Enzyme and Microbial Technology, January 2004, vol. 34, no. 1, p. 33-40. [CrossRef] KONDO, A.; LIU, Y.; FURUTA, M.; FUJITA, Y.; MATSUMOTO, T. and FUKUDA, H. Preparation of high activity whole cell biocatalyst by permeabilization of recombinant flocculent yeast with alcohol. Enzyme and Microbial Technology, December 2000, vol. 27, no. 10, p. 806-811. [CrossRef] KONSOULA, Z. and LIAKOPOULOU-KYRIAKIDES, M. Co-production of α-amylase and β-galactosidase by Bacillus subtilis in complex organic substrates. Bioresource Technology, January 2007, vol. 98, no. 1, p. 150-157. [CrossRef] LIU, D.L.; YAO, D.S.; LIANG, Y.Q.; ZHOU, T.H.; SONG, Y.P.; ZHAO, L. and MA, L. Production, purification, and characterization of an intracellular aflatoxin-detoxifizyme from Armillariella tabescens (E-20). Food and Chemical Toxicology, May 2001, vol. 39, no. 5, p. 461-466. [CrossRef] LIU, C.H.; LU, W.B. and CHANG, J.S. Optimizing lipase of Burkholderia sp. by response surface methodology. Process Biochemistry, September 2006, vol. 41, no. 9, p. 1940-1944. [CrossRef] MARTINS, D.B.G.; SOUZA, C.G.; SIMÕES, D.A. and MORAIS, M.A. The β-galactosidase activity in Kluyveromyces marxianus CBS6556 decreases by high concentrations of galactose. Current Microbiology, May 2002, vol. 44, no. 5, p. 379-382. [CrossRef] MILLER, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry, 1959, vol.31, no. 3, p. 426-428. [CrossRef] MYERS, R.H. and MONTGOMERY, D.C. Response surface methodology: Process and product optimization using designed experiments. John Wiley & Sons, Inc, New York, 1995. 705 p. ISBN 0-471-58100-3. NAESSENS, M.; VERCAUTEREN, R. and VANDAMME, E.J. Three-factor response surface optimization of the production of intracellular dextran dextrinase by Gluconobacter oxydans. Process Biochemistry, June 2004, vol. 39, no. 10, p. 1299-1304. [CrossRef] NAWANI, N.N. and KAPADNIS, B.P. Optimization of chitinase production using statistics based experimental designs. Process Biochemistry, February 2005, vol. 40, no. 2, p. 651-660. [CrossRef] NUMANOGLU, Y. and SUNGUR, S. β-galactosidase from Kluyveromyces lactis cell disruption and enzyme immobilization using a cellulose-gelatin carrier system. Process Biochemistry, February 2004, vol. 39, no. 6, p. 705-711. [CrossRef] PANESAR, R.; PANESAR, P.S.; SINGH, R.S.; KENNEDY, J.F. and BERA, M.B. Production of lactose-hydrolyzed milk using ethanol permeabilized yeast cells. Food Chemistry, 2007, vol. 101, no. 2, p. 786-790. [CrossRef] PARK, P.K.; KIM, E.Y. and CHU, K.H. Chemical disruption of yeast cells for the isolation of carotenoid pigments. Separation and Purification Technology, February 2007, vol. 53, no. 2, p. 148-152. [CrossRef] RAJOKA, M.I.; KHAN, S. and SHAHID, R. Kinetics and regulation studies of the production of β-galactosidase from Kluyveromyces marxianus grown on different substrates. Food Technology and Biotechnology, October-December 2003, vol. 41, no. 4, p. 315-320. RECH, R.; CASSINI, C.F.; SECCHI, A. and AYUB, M.A.Z. Utilization of protein-hydrolyzed cheese whey for production of β-galactosidase by Kluyveromyces marxianus. Journal of Industrial Microbiology & Biotechnology, August 1999, vol. 23, no. 2, p. 91-96. [CrossRef] SANTOS, A.; LADERO, M. and GARCÍA-OCHOA, F. Kinetic modeling of lactose hydrolysis by a β-galactosidase from Kluyveromyces fragilis. Enzyme and Microbial Technology, May 1998, vol. 22, no. 7, p. 558-567. [CrossRef] SCHNEIDER, A.L.S.; MERKLE, R.; CARVALHO-JONAS, M.F.; JONAS, R. and FURLAN, S. Oxygen transfer on β-D-galactosidase production by Kluyveromyces marxianus using sugar cane molasses as carbon source. Biotechnology Letters, April 2001, vol. 23, no. 7, p. 547-550. [CrossRef] SHAIKH, S.A.; KHIRE, J.M. and KHAN, M.I. Production of β-galactosidase from thermophilic fungus Rhizomucor sp. Journal of Industrial Microbiology & Biotechnology, October 1997, vol. 19, no. 4, p. 239-245. [CrossRef] SONG, D.D. and JACQUES, N.A. Cell disruption of Escherichia coli by glass bead stirring for the recovery of recombinant proteins. Analytical Biochemistry, June 1997, vol. 248, no. 2, p. 300-301. [CrossRef] TARI, C.; GÖGUS, N. and TOKATLI, F. Optimization of biomass, pellet size and polygalacturonase production byAspergillus sojae ATCC 20235 using response surface methodology. Enzyme and Microbial Technology, April 2007, vol. 40, no. 5, p. 1108-1116. [CrossRef] UMA MAHESWAR RAO, J.L. and SATYANARAYANA, T. Improving production of hyperthermostable and high maltose-forming α-amylase by an extreme thermophile Geobacillus thermoleovorans using response surface methodology and its applications. Bioresource Technology, January 2007, vol. 98, no. 2, p. 345-352. [CrossRef] Note: Electronic Journal of Biotechnology is not responsible if on-line references cited on manuscripts are not available any more after the date of publication. |